
Anatomy Of Flowering Plants Introduction
Anatomy Of Flowering Plants The living world shows diversity in terms of organisms’ external and internal features. Numerous scientists have descriptions of different organisms based on observations made through the naked eye and/or under the microscope.
The branch of science dealing with the internal structure and organisation of organisms is called anatomy (Greek Ana asunder or into pieces and temnein to cut).
One of the branches of anatomy is histology (Greek Histos tissue and logia= knowledge or study), which includes a study of cellular arrangements into a tissue or higher level of organisation and how such organisation forms an organism.
Like all organisms, plants are also made up of tissues. All these tissues organise together to form vegetative organs of the plants, such as roots, stems, leaves, etc. In this chapter, we shall learn how the different types of tissues originated and organised to form the different organs in plants.
Read and Learn More: WBCHSE Notes for Class 11 Biology
Tissue
Tissue Definition: A tissue is a collection of cells of the same origin and has the same methods of development, performing a specific function in a harmonious way.
The cell is the structural and functional unit of a living organism. All organisms are formed of either a single or a group of cells. For a single-celled organism, all of its biological activities are accomplished within the same cell. In the case of multicellular organisms, the cells are organised into tissues which perform specific functions.
| Class 11 Biology | Class 11 Chemistry |
| Class 11 Chemistry | Class 11 Physics |
| Class 11 Biology MCQs | Class 11 Physics MCQs |
| Class 11 Biology | Class 11 Physics Notes |
Again, different tissues are organised to form a tissue system. Several tissue systems together form an organ and several organs collectively perform specific physiological activities for the whole organism.
Cell -> Tissue -> Tissue system -> Organ -> Organism
Short notes on anatomy of flowering plants for Class 11
Plant Tissue- Meristematic And Permanent Tissue
The plant tissues may be classified on the basis of different characteristics viz., their position in the plant body, types of constituting cells, functions, the methods of development and origin, etc. However, on the basis of origin and stages of development, the tissues are grouped into
- Meristematic tissue and
- Permanent tissue.
Meristematic Tissue
Meristematic Tissue Definition: Meristematic tissue or meristem is defined as the tissue, in which the cells continuously divide for an indefinite period to add new cells to the plant body.
In the early embryonic stage, all the cells of the embryo remain actively divisible. But, as the embryo develops into a seedling the dividing property of the cells becomes restricted to some specific regions or zones.
The tissues at these zones are called meristems or meristematic tissues. the word meristem has been derived from the Greek word meristos which means divisible.

Meristematic Tissue Characteristics:
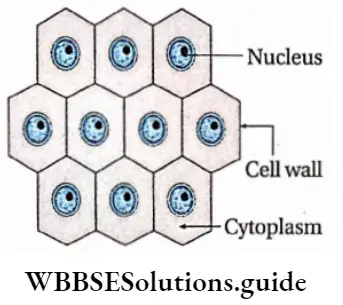
- Meristematic cells are living, undifferentiated (not determined to form any specific tissue), isodiametric (having equal diameter) and are usually small and without any intercellular spaces.
- Each cell possesses one large and prominent nucleus, and dense cytoplasm with or without small scattered vacuoles, known as pro-vacuoles.
- Cells are spherical, oval or polyhedral.
- The cell wall is thin, homogeneous and composed of cellulose.
- Cells contain proplastids (precursor of plastids), poorly developed endoplasmic reticulums, and mitochondria with fewer cristae. Cells lack ergastic substances.
- Cells are capable of dividing for indefinite periods. Meristematic cells which remain active throughout their lifespan are called initiating cells and the cells derived from them are called derivatives.
- The rate of respiration is high in meristematic cells. So, the amount of stored food is scanty.
Meristematic Tissue Function:
- Meristems divide continuously to increase the number of cells in the plant body.
- This brings about the growth and development of the plant body as a whole through different tissue and organ formation.
- The derivatives generated from initiating cells gradually enlarge, change their shape, and ultimately mature with definite shapes and perform specialised functions. They further mature to form permanent tissues. This process of maturation is referred to as differentiation.
- These tissues are responsible for the formation of branches, leaves and flowers.
- New vascular tissues, which take part in the transportation of water, minerals and food, are formed from specified meristematic tissue called vascular cambium. New vascular tissues bundle up at the core of the plant to develop vascular bundles. Further action of this meristem is to increase the girth of the plants.
- Cork is produced from cork cambium to protect the internal structures of the stem and root.
- They help to form root hairs too.
Meristematic Tissue Distribution: Meristems are distributed mainly at the growing tips. These regions include the main and lateral shoot apices, root apices, bases of internodes, flower buds and leaf apices.
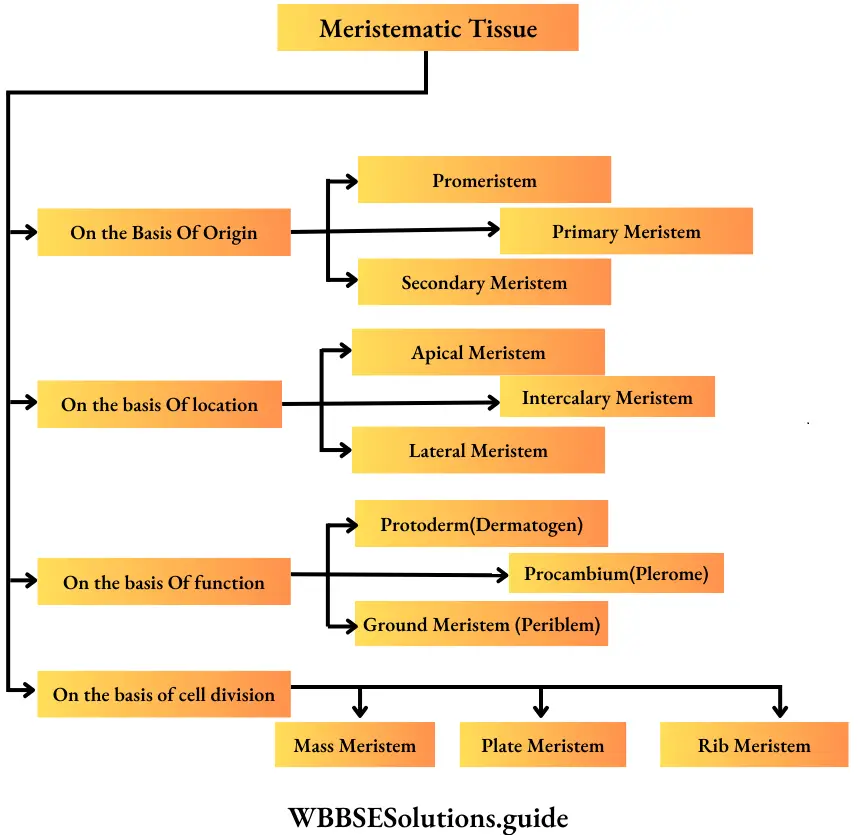
Meristematic Tissue Classification: Meristematic tissues were classified on the basis of origin and development, location in the plant body, function and plane of division.
Classification of meristem based on origin
On the basis of origin and development, meristems are of the following types—
Promeristem or Primordial meristem
Primordial meristem Definition: The tissue which is present at the tip of growing regions of a plant right from its embryonic stage, is known as primordial meristem.
Primordial meristem Location: Stem and root apices.
Primordial meristem Characteristics:
- This meristem develops from embryo cells and is also known as embryonic meristem.
- Cells are small, immature and lack vacuoles. The cell wall is very thin.
- Intercellular spaces are absent between the cells.
Primordial meristem Function: This tissue divides continuously to form primary meristem, which initiates the formation of new plant parts.
Promeristem -> Primarymeristem -> Apical meristem

Primary Meristem
Primary Meristem Definition: The tissues that originate directly from the embryonic meristem and retain meristematic activity throughout their life span, are called primary meristem.
Primary Meristem Location: Root, stem and leaf apices. Also, present between the internode.
Primary Meristem Characteristic:
- Primary meristem develops from the primordial meristem.
- This meristem develops in the plants at the embryonic stage and continues to divide throughout the lifespan of the plant.
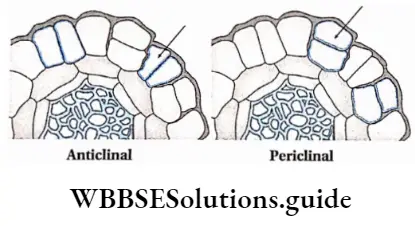
- Cells of this tissue divide anticlinally or periclinally.
Types of cell division on the basis of divisional plane
Anticlinal cell division: The type of cell division where the plane of division is at the right angle to the surface of the plant body is known as anticlinal cell division.
Periclinal cell division: The type of cell division where the plane of division is parallel to the surface of the plant body is known as periclinal division.
Primary Meristem Function:
- Primary parts of the plant are produced from the primary meristems. Intrafascicular or fascicular cambium produce secondary vascular tissues.
- Cells of the primary tissue divide only in one plane and convert into immature permanent tissues.
- These immature permanent tissues can no longer divide or develop to form different permanent tissues. These tissues are known as primary permanent tissue.
- A group of different types of permanent tissues together forms a tissue system. The tissue systems differentiate and give rise to primary bodies i.e., root, stem and leaf.
- This tissue causes the primary growth of the plants.
Secondary meristems
Secondary meristems Definition: Secondary meristems are those meristematic tissues that develop from permanent tissues after they regain their ability to divide.
Secondary meristems Location: This tissue is found in the mature regions with secondary growth of the plant. This type of tissue is known as cambium.
Secondary meristems Characteristics:
- The cells of secondary meristematic tissues have vacuoles and thick cell walls.
- Cells in these tissues contain a large nucleus and dense cytoplasm.
- The cells also contain secretory substances and excretory products.
Secondary meristems Functions:
- The secondary meristems add new cells to the primary body forming supplementary tissues during secondary growth.
- It thickens the bark and increases the breadth of the tree.
- It also gives protection and helps to repair wounds.
- Cells of the secondary meristem divide to form secondary permanent tissue. The interfascicular cambium produces secondary xylem, secondary phloem, conjunctive tissues and medullary rays (radially arranged parenchymal cells between two vascular fascicles).
- Secondary meristem gives rise to secondary tissue for wound healing.
Secondary meristems Types: There are different types of secondary meristems
Interfascicular cambium: These have originated from primary medullary rays (a primary tissue, extending between vascular bundles). This type of cambium is located between two vascular bundles.
Cork cambium or phellogen: These tissues have originated from the hypodermis, epidermis and outermost layer of cortex (cell layers between epidermis and endodermis). It forms the phellem or cork at the outer side and the phelloderm at the inner side. The phellogen, phellem and phelloderm together are known as periderm.
Wound cambium: This tissue originates injured part and heals the wound.
Accessory cambium: These are present in the lower region of the phloem. In monocotyledonous plants, cambium is generally absent. But in plants, this tissue may be found, then it is called an accessory cambium. These tissues are responsible for abnormal secondary growth in monocot plants, like Dracena, etc.
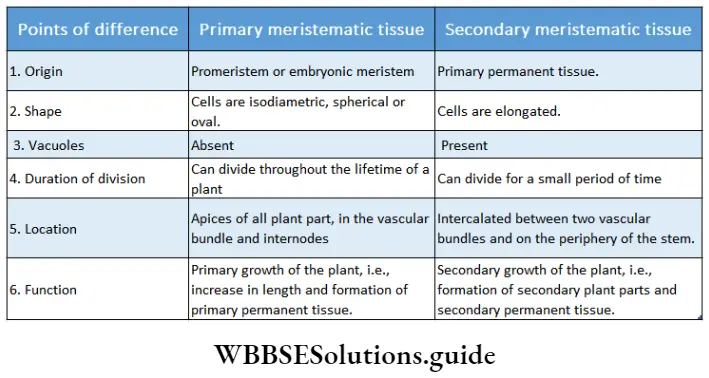
Classification of meristem on the basis of location
On the basis of location, there are three types of meristems—apical meristem, lateral meristem and intercalary meristem.
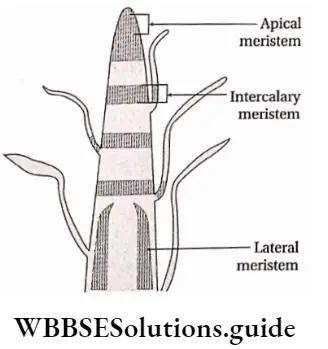
Apical meristem
Apical meristem Definition: The meristem which is found at the shoot and root apices of the main and lateral branches is called apical meristem.
Apical meristem Location: Root and shoot apices. Apical meristem includes the pro meristem and primary meristem.
Apical meristem Characteristics:
- Cells of the apical meristem are known as apical cells. These cells are always in the terminal (at the shoot apex) or subterminal, i.e., just below the outermost layer (in the root apex).
- A single apical cell is found in the apical meristem of the lower group of plants (algae, bryophytes and pteridophytes); but in the case of a higher group of plants (gymnosperms and angiosperms) a group of cells constitute the apical meristem, called apical initials.
- This meristem is also known as a growing point as its activity results in plant growth.
- It is the origin of primary permanent meristem
Apical meristem Function:
- The increase in length of the plant axis is mainly achieved by the apical meristems.
- By continuous division, these tissues give rise to permanent tissue. These permanent tissues together form different parts of the plant.
- Leaves grow due to the activation of the apical meristem of the shoot apex.
Apical meristem Structural development of apical meristem: Different structural developments are found mainly in the root and shoot apical meristems that are found in the root and shoot apices respectively.
Shoot apex and root apex are discussed under separate heads.
Root apex
Root apex Definition: The root tip that remains protected by the root cap arid contains clusters of primary cells, is known as the root apex.
Root apex Characteristics:
- This portion is derived from the radicle of the embryo.
- Root apex does not contain branch primordia and leaf primordia.
- Meristematic tissues are present in subterminal regions due to the presence of root cap and calyptra.
- The root apex does not show any periodic changes in shape and structure.
- Root apex not only produces cells towards the axis but also away from it.
Theories related to the structural organisation of root apex: To understand the structure and activity of root apical meristem various theories were proposed by scientists. Some of those related to the structure and activity of root apical meristem are discussed below.
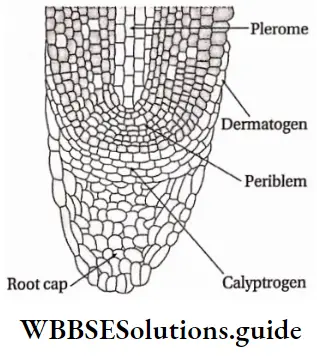
Root apex Histogen theory: In his histogen theory of shoot apex, Hanstein (1868) also included root apex.
The theory explains that the group of primary meristems (histogen) are divided into three regions—
- The dermatogen region forms the root epidermis (epiblema) and the root cap (in dicotyledons).
- The endodermis and the cortex are formed by the periblem.
- The plerome region gives rise to pericycle, central vascular tissues and pith. This theory suggests that the root cap is derived from a separate group of cells, called calyptrogen.

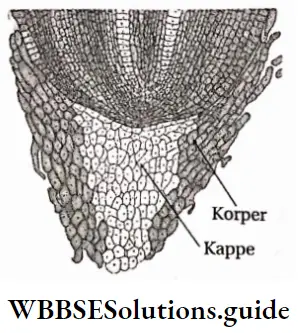
Korper-Kappe theory: This theory was proposed by scientist Schuepp (1917). According to this theory, the cells of the root cap divide to form Korper and Kappe regions. Characteristic cell division is found in these regions. Cells towards the periphery of the root divide transversely to form Kappe cells and inner cells divide longitudinally to form Korper cells.
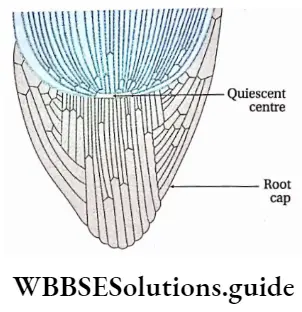
Quiescent centre
Scientist Clowes (1961) named the group of inactive cells, present in the form of a hemisphere or disc, between the root cap and active meristem as a quiescent centre. It is found just behind the root cap region in Zea mays.
Characteristics of quiescent centre:
- Cells of this region either remain in G0 phase or show a very slow rate of mitotic division.
- The rate of DNA and protein synthesis is also very slow in this region.
- This region is the centre of the root apex.
- Cells below the quiescent centre are active and give rise to the root cap.
Lateral meristem
Lateral meristem Definition: The laterally situated meristems which are parallel to the surface of the plant body and are composed of a single layer of rectangular cells that produce secondary permanent tissues are known as lateral meristems.
Lateral meristem Location: These tissues run parallel to the axis of the root and stem.
Lateral meristem Characteristics:
- Cells of these tissues divide periclinal to produce secondary permanent tissue
- Lateral meristem includes both primary and secondary meristem.
- The cells of this tissue are rectangular and arranged in a single layer.
- The fascicular cambium and the phellogen or cork cambium are examples of this type of meristem.
Lateral meristem Function: By the activity of the lateral meristem secondary growth occurs in the plant. The activity of lateral meristem develops cork, heals wound and the plant body increases in girth or diameter.
Intercalary meristem
Intercalary meristem Definition: The meristems that are located between the regions of permanent tissues during the development of apical meristems are known as intercalary meristems.
Intercalary meristem Location: Intercalary meristems are found in different organs of plants, such as the leaf bases in Pinus, internode bases in the stems of grasses and Equisetum and at the base of the node as in Mentha sp., etc.
Intercalary meristem Characteristics:
- Cells of these tissues are elongated but their structures are similar to the primary meristems.
- These tissues are found along the axis of the plants.
- The life span of the intercalary meristem is short as they get converted into permanent tissues after a short period of time.
Intercalary meristem Function:
- The main axis and its branches increase in length by the activity of this type of meristem.
- It increases the length of the internodes.
- It also increases the length of the leaf base.

Classification of meristem based on function
After the development of pro meristems in the embryonic stage, they give rise to primary meristems. These meristems generate all types of tissues in plants.
Thus, when scientists categorised meristems functionally, they based their decisions on the functions of different layers of apical meristems of root and shoot.
The different types of meristems according to their functions are as follows—
Types of meristem according to Haberlandt:
Haberlandt classified the meristem into 3 types according to their functions. This was based on his work on apical meristem.
They are as follows—
Protoderm
Protoderm Definition: The outermost cell layer of the primary meristem that gives rise to the epidermis by periclinal division is called the protoderm.
1. Protoderm Characteristics:
- It is the outermost layer of the primary meristem.
- The cells of this meristem divide periclinal to form the epidermis in root and shoot,
- This causes the growth of the dorsal region of the different plant parts and also gives rise to epidermal hairs.
2. Protoderm Function:
- Protoderm gives rise to epiblema and epidermis,
- It also gives rise to epidermal hairs and epidermal cells.
Procambium
Procambium Definition: The elongated tapering cells, present at the centre of the apical meristem and giving rise to vascular bundle are known as procambium.
Procambium Characteristics:
- The elongated and tapering cells present in clusters at the growing region are called procambial strands,
- They form a ring in the case of dicot stems and remain scattered in monocot stems,
- The procambial strands give rise to vascular bundles, consisting of the primary xylem towards the centre and the primary phloem towards the periphery.
- A single procambial strand is present at the centre of the root,
- The procambium strands give rise to pericycle in some stems.
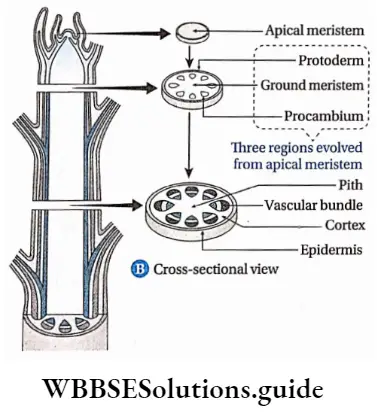
Procambium Function:
- It forms vascular bundles in the roots which are radially arranged,
- The vascular bundle of the stem originates from the procambium. These vascular bundles of stem may be open or closed type.
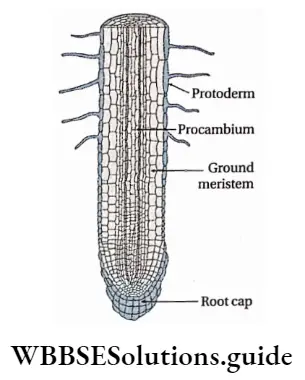
Fundamental or Ground meristem
Ground meristem Definition: The part of the primary meristematic tissue other than the procambium and protoderm, is known as ground meristem.
1. Ground meristem Characteristics:
- Cells of these tissues undergo anticlinal or peridinal division to form primary meristems.
- This tissue is present inside and outside the stellar regions.
2. Ground meristem Function: It forms the hypodermis, cortex, medullary rays, and endodermis, outside the stellar regions. It develops the pericycle and the pith inside the stele.
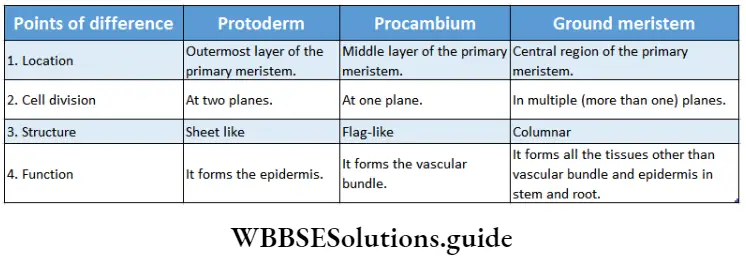
Types of meristem according to Hanstein: Hanstein divided meristem into three categories on the basis of their functions—dermatogen, periblem and plerome. This was based on ‘Histogen theory’, on apical meristem, proposed by him in 1868.
Dermatogen
Dermatogen Definition: The outermost layer of the primary meristematic tissue, that gives rise to protoderm is called dermatogen.
Dermatogen Characteristics: Cells divide anticlinally.
2. Dermatogen Function: The epidermis and epiblema are formed by the cells produced by the anticlinal division.
Periblem
Periblem Definition: The layer of primary meristem between the dermatogen and plerome from which components of ground meristem are formed is known as periblem.
Periblem Characteristics: Cells of this layer can divide both anticlinally and periclinally.
Periblem Function: It forms different parts of extrastellar regions (hypodermis, cortex, endodermis) and intrastellar regions (pericycle and pith).
Pierome
Pierome Definition: The innermost layer of the primary meristematic tissue, that gives rise to stele orprocambium is known as plerome.
1. Pierome Characteristics: Cells of this tissue divide periclinal.
2. Pierome Function: It gives rise to stele, by periclinal division
Classification of meristem based on plane of cell division
Based on the plane of cell division, meristems are of three types—mass meristem, plate meristem and rib meristem.
Mass meristem
Mass meristem Definition: The meristem, that divides in all planes and produces an irregular mass of cells, is known as mass meristem.
Mass meristem Characteristics: These cells divide in all planes resulting increase in the volume of the plant body.
Mass meristem Function: Mass meristems give rise to the cortex and pith. This tissue also takes part in the early stages of the development of the embryo, endosperm, sporangia, etc.
Plate meristem
Plate meristem Definition: The meristem, whose cells undergo anticlinal division in two planes and cause a plate-like increase in the surface area of plant parts is known as plate meristem.
Plate meristem Characteristics:
- These cells divide anticlinally.
- Cells are flat and distributed only in one plane.
- This tissue can be uniseriate or multiseriate. However, the number of cell layers does not increase with a further increase in cell number. Thus, it grows in the surface area.
Plate meristem Function:
- Single cell-layered plate meristem forms the epidermis.
- Several cell-layered thick plate meristem is responsible for the development of leaf blade.
Rib meristem
Rib meristem Definition: The meristem whose cells form columns or rows of cells by repeated anticlinal divisions in one plane is called rib meristem.
Rib meristem Characteristics: Cells are linearly arranged due to anticlinal division. As a result, the cells look like ribs.
Rib meristem Function: This meristem becomes active during the formation of young roots, cortex and pith in young stems. It also helps in the formation of algal filaments, etc.
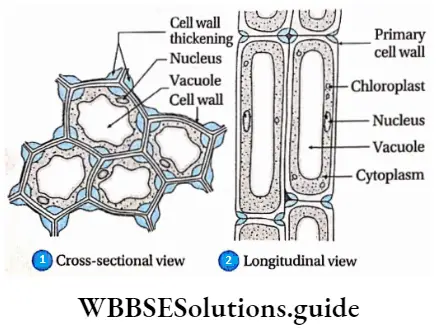
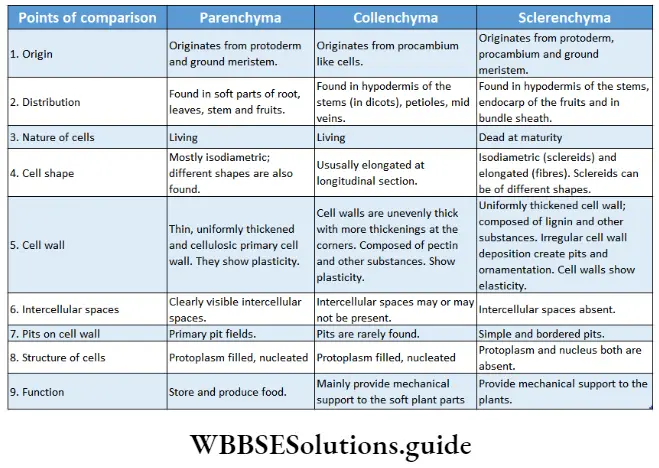
Collenchyma or Collocate
Collenchyma or Collocate Definition: Collenchyma or collocate is a type of primary, permanent simple tissue consisting of elongated living cells with uneven cellulosic cell walls and angular thickening.
Collenchyma or Collocate Origin: Originates from certain elongated cells resembling procambium, formed in the very early stages of differentiation of the meristem.
Collenchyma or Collocate Distribution: These tissues are found as supporting cells or mechanical tissues in the soft mature parts of the plants. These tissues are found in young leaves, stems and petioles. They are uniformly or non-uniformly distributed just below the epidermis (hypodermis) of dicotyledonous plants.
Collenchyma or Collocate Characteristics:
- Younger collenchyma cells show more extensibility and plasticity than the older ones.
- Usually, collenchyma cells are polygonal in cross-section.
- These are living cells with large vacuoles.
- Cell walls are unevenly thickened. Deposition of cell wall material is higher at the corners of the cells.
- Collenchyma cells vary in size and shape. The smaller cells resemble parenchyma cells. The older and longer cells resemble fibres as they have overlapping tapering ends.
- Cells of this tissue may contain chloroplasts and carry out photosynthesis.
- In some cases, collenchyma cells store tannins as secondary metabolites.
- The cell wall consists of cellulose, a high amount of hemicellulose, and pectic materials. However, lignin is completely absent.
- The collenchyma cells can undergo reversible changes and regain the divisional property.
- Primary pit fields can be distinguished in the walls of collenchyma cells.
- Collenchyma cells may or may not have intercellular spaces. Often intercellular space is filled with cell wall materials.
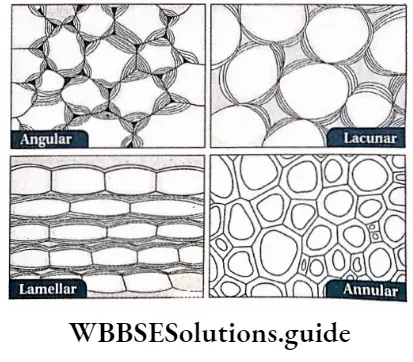
Collenchyma or Collocate Types: According to cell wall thickening, four main types of collenchyma are recognised.
They are as follows—
- Angular collenchyma: In these cells, the cell wall material depositions or thickening are localised at the corners or angles of the cells to form a compact tissue. It is found in the stems of Datura, Dahlia, Cucurbita, Solanum tuberosum, Atropa belladonna, etc., and in the petioles of the leaves of Vitis, Begonia, Coleus, Cucurbita, Beta, Morus, etc.
- Lacunar collenchyma: In these cells, thickenings appear around the intercellular spaces. This type of collenchyma is also called tubular collenchyma. It is found in the leaf petioles of Salvia, Malva, Althaea, Asclepias and in the members of Compositae.
- Plate or Lamellar collenchyma: In this type of collenchyma, cells are compactly arranged without intercellular spaces. Thickenings occur in various patterns mainly on the tangential walls of the cells. This type of collenchyma tissue is found in stems of Sambucus nigra and Rhamnus, etc., and in the petiole of Cochlearia armoracia.
- Annular collenchyma: In this type of tissue, the cell wall materials are uniformly deposited towards the centre, which provides a ring-like structure to the cells. Angular collenchyma sometimes transforms into annular collenchyma due to the uniformity of deposition. This collenchyma is found in carrot leaves.
Collenchyma or Collocate Functions:
- It functions as the supporting tissue.
- Collenchyma cells give protection and mechanical support to the growing plant parts.
- These cells impart flexibility and elasticity to the plant parts,
- Photosynthesis takes place in the chloroplast containing collenchyma cells.
- It can also store food.
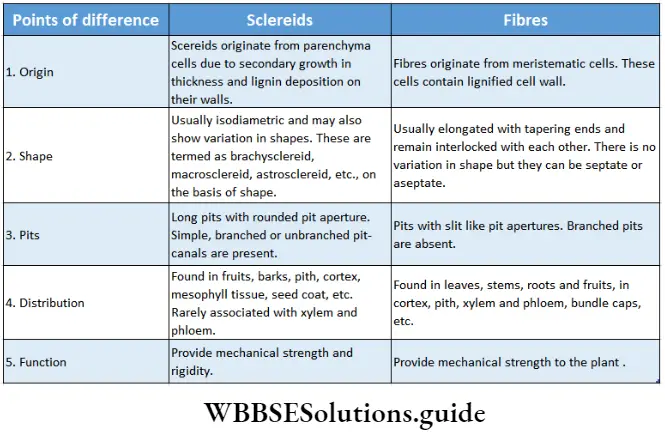
Sderenchyma
Sderenchyma Definition: The tissue, composed of elongated dead cells with very thick, hard and lignified secondary walls and without any intercellular spaces is called sclerenchyma tissue.
Sderenchyma Origin: Originates from protoderm, ground meristem and procambium.
Sderenchyma Distribution: Present in the pericycle, bundle cap, hypodermis, etc. This tissue is also present in the seed coat of peas, green beans, etc., and the endocarp of apples, etc.
Sderenchyma Characteristics:
- Cells of the sclerenchyma tissue differ in shape, structure, origin and development. Cells can be tapered, elongated, star-shaped or oval in shape,
- Mature cells of the sclerenchyma tissue are dead with an almost obliterated cell cavity or lumen.
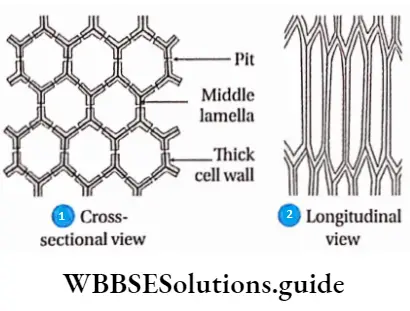
- Several unthickened or non-lignified areas called simple pits were found. Sometimes these pits have a border or rim of cell wall materials, thus called bordered pits.
- The thick cell wall is composed of cellulose, hemicellulose and lignin.
- Cell wall materials are even deposited inside the cell lumen and intercellular spaces.
Sderenchyma Types: According to the shape and size, sclerenchyma is of two types—sclerenchyma fibre and steroids or sclerotic cells. These are discussed under separate heads below.
1. Sclerenchyma fibre: These are much elongated and narrow, spindle-shaped cells with tapered ends.
- Sderenchyma Origin: Fibres originate from meristematic cells of protoderm or ground meristem.
- Sderenchyma Distribution: Fibres remain distributed in different organs of the plant body. In the leaflets of Cycas, they occur singly as idioblasts. They may occur in separate strands in the cortex or as sclerenchymatous patches or bundle caps above the vascular bundles or in the vascular bundle as components of the xylem and the phloem.
- Sderenchyma Characteristics:
-
- Cell walls are uniformly thickened and highly lignified with simple pits.
- Cell lumen is reduced due to much thickened secondary wall and deposition of cell wall materials inside the lumen,
- The fibres are always dead at maturity,
- They appear polygonal in cross-sectional view and elongated and tapered at both ends in the longitudinal view,
- In certain cases, the fibre walls are cellulosic and non-lignified. Some fibres have mucilaginous walls.
- These fibres remain overlapped over one another.

4. Sderenchyma Types: Based on the positions in the plant body, fibres are classified into different types
- Xylary fibres or wood fibres refer mainly to the sclerenchyma fibres that are associated with the xylem.
- Extraxylary fibres refer to the sclerenchyma fibres present at the outermost surface of the xylem. These are also known as bast fibres.
- Surface fibres are present on the outer surface of the fruits and seeds. Based on the cell wall properties and amount of pits xylary fibres are again divided into three types— libriform fibres, fibre tracheids and mucilage fibres.
5. Sderenchyma Function: These tissues are present in woody and fibrous parts of the plants and provide mechanical support. Jute fibres, coconut fibres, etc., are examples of sclerenchyma fibres.
Three types of wood fibres
- Libriform fibres: The cell wall of this type of xylem fibre is thick, and contains simple pits. Cells are of medium length.
- Fibre-tracheids: The cell wall of this type of xylem fibre is thin, with a bordered pit. The cells are elongated.
- Mucilaginous or gelatinous fibres: The cell wall of this type of xylem fibres is mucilaginous or gelatinous.
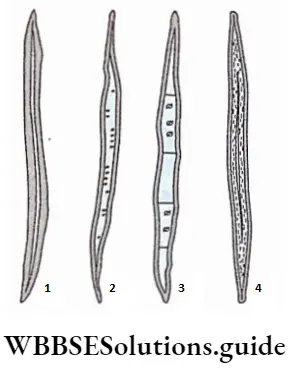
2. Sclereids or sclerotic cells: Sclereids or sclerotic cells are short isodiametric or irregularly shaped cells with pit canals that die at maturity.
- Sderenchyma Origin: Sclereids originate due to the secondary thickening of the cell walls of the parenchymatous cells. The secondary wall becomes thickly deposited in numerous concentric layers with the formation of simple pits that contain branched or unbranched pit canals. The mode of development of all types of sclereids is common but the number of pit formations varies.
- Sderenchyma Distribution: Sclereids are abundantly present in the cortex, phloem, pith, mesophyll tissue, etc., as isolated individual cells or in clusters. They are found in the outermost covering of fruit or the pericarp of Pyrus, Psidium, etc. They also occur in the hard innermost covering or endocarp and seed coats of many plants either singly or in clusters. They are the major components in the shells of walnuts and seed coats of peas.
- Sderenchyma Characteristics:
- Cells of this tissue are of different shapes and sizes,
- Cells are columnar, elongated, star-shaped, etc.
- The cell wall is thick and composed of cellulose, and hemicellulose and also has a high amount of lignin, suberin and cutin.
- The sclereid walls possess unbranched simple pits or simple pits with branched pit canals,
- Some of the isodiametric, lignified, hard and thick-walled sclereids are called stone cells.
- They remain as hard mosaics of cells intermingled with soft parenchyma in different places of the plant body.
- In many cases, sclereids may appear as idioblast and are found to occur in the inter-cellular spaces.
4. Sderenchyma Types: According to the shape, size and nature of wall thickening
The sclereids are categorised into the following types—
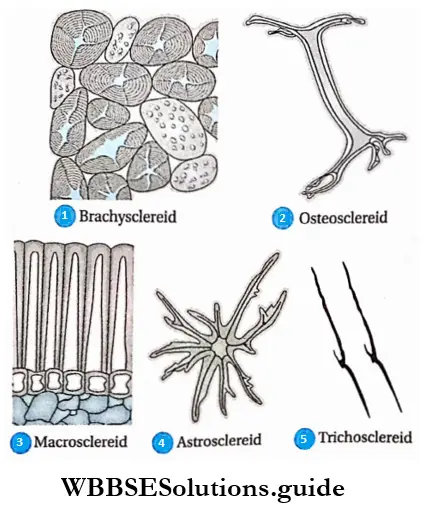
- Brachysclereids or stone cells, these sclereids are more or less isodiametric in appearance. They are also called grit cells, as they provide a gritty texture to the pulp of many fruits like Pyrus sp., Psidium sp., etc. Brachysclereids are usually distributed in the phloem, the cortex and the bark of stems.
- Macrosclereids or rod cells, are rod-shaped columnar sclereids which often form a continuous palisade parenchyma-like epidermal layer in the outer seed coat or testa of leguminous seeds. Macrosclereids occur in the pulp of Malus sylvestris (apple).
- Osteosclereids are elongated bone or spool-shaped sclereids which remain in columnar arrangement. The ends of these sclereids are enlarged, lobed, or sometimes branched. Such sclereids are mainly found in seed coats and leaves of certain dicotyledons like Pisum sp., Hakeo sp., etc.
- Astrosclereids, are branched and often star-shaped in appearance. They are mainly found in leaves and stems of many dicotyledonous plants like Thea sp., and Nymphaea sp. Trochodendron sp., etc.
- Trichosclereids, this type of sclereids are very elongated, hair-like, branched sclereids. They are found in the intercellular spaces in the leaves and also in the stems and aerial roots of certain plants.
5. Sderenchyma Function:
- Provide stiffness to the part they occur,
- Form seed coat in leguminous plants.
- Provide mechanical strength to the endocarp in some fruits.
- Protect the plants from adverse weather conditions.
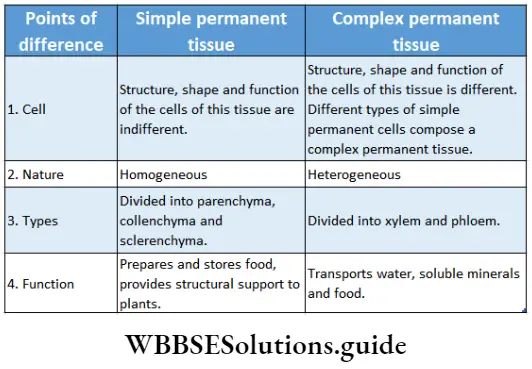
Complex permanent tissues
Complex permanent tissues Definition: Complex permanent tissues are composed of two or more types of simple tissues and are heterogeneous in nature.
Complex permanent tissues Characteristics:
- Complex permanent tissues are composed of two or more types of simple permanent tissues. Thus cells of this type of tissue can be of different shapes and sizes.
- This tissue is formed of different components of a single meristematic tissue.
- Different cell components together perform one special function.
- Cells of complex permanent tissues can be living or dead.
Complex permanent tissues Types: Complex permanent tissue is mainly of two types—xylem and phloem. They together comprise the vascular tissue system of a plant. Xylem and phloem are discussed below in separate heads.
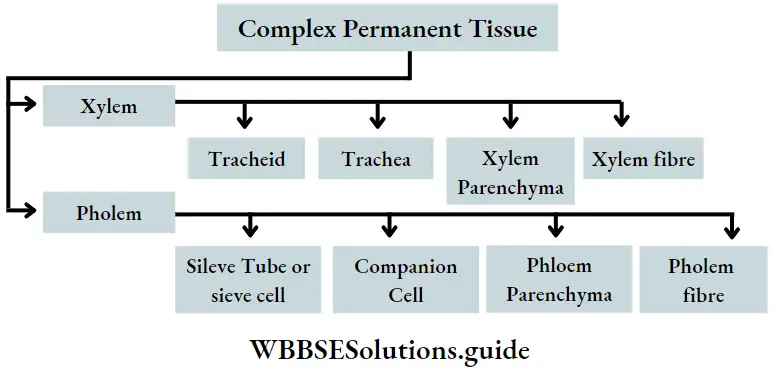
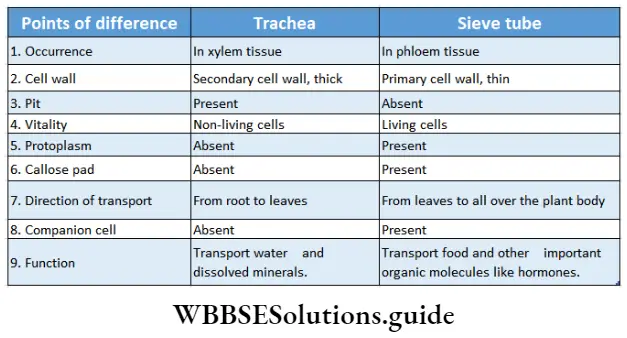
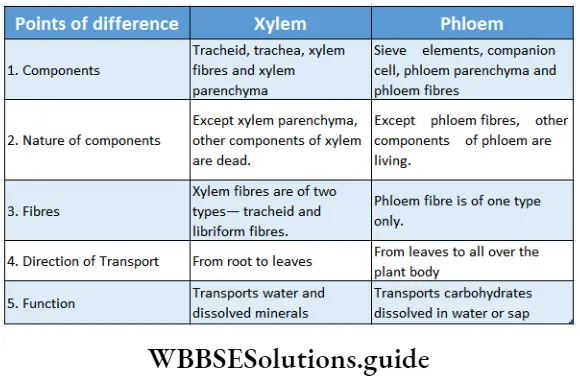
Xylem
Xylem Definition: Xylem is the complex permanent tissue that comprises a part of the vascular system and helps in the conduction of water from the root.
Xylem Origin: The primary xylem is derived from the procambium, whereas the secondary xylem is derived from the vascular cambium(fascicular and interfascicular I cambium together)during secondary growth.
Xylem Distribution: In flowering plants, found in the j vascular bundles of root, leaves and stems. Xylem is also present in the root and stem of the pteridophytes.
Xylem Function:
- The main function is the circulation: of water and dissolved minerals from the xylem of root I to the same of the leaves.
- It provides mechanical strength to the plants.
- It stores produced food and waste products.
Xylem Components: Depending on the origin, the xylem is of two types— primary and secondary. This complex tissue is composed of both living and non-living cells.
The main components of the tissue are—
- Tracheids
- Tracheae or vessels,
- Xylem parenchyma and
- Xylary fibres or wood fibres. Tracheids and tracheae together are known as tracheary elements. These components are discussed below in separate heads.
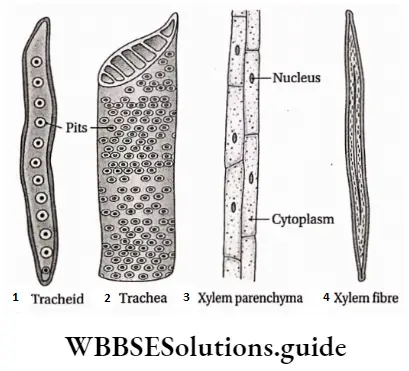
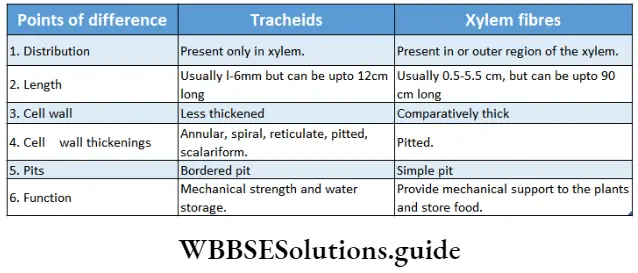
Tracheids: The tracheids are dead, elongated, lignified thick-walled cells with narrow ends.
Xylem Origin: In the primary xylem tracheids originate from procambium and in the secondary xylem, they originate from the cambium ring from a single fusiform (tapered at both ends) initial.
Xylem Distribution: Tracheids are found in the primary and secondary xylems of vascular plants. They predominantly occur in pteridophytes, gymnosperms and primitive angiosperms.
Xylem Structure:
- Tracheids remain parallel to the long axis of the plant part, where they occur.
- The ends may also be blunt, rounded, chisel-like or oblique.
- They are dead with larger cell lumen.
- The hard and lignified cell wall contains bordered pits.
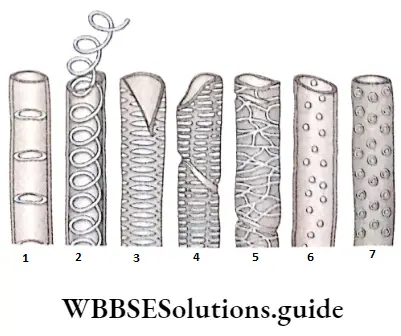
- They possess various kinds of wall thickening or ornamentations like annular, spiral, scalariform and reticulate thickening.
- In cross-section, these cells appear angular, polyhedral or round in outline.
- These cells remain one above the other with overlapping ends.
- The transverse walls have many perforations.
- Communication with surrounding cells is established through bordered pits on the lateral walls of adjacent tracheids.
Xylem Functions:
- The primary function of the tracheid is the conduction of water and dissolved minerals in it.
- It also provides mechanical support to plants.
- Tracheids also store water in some plants.
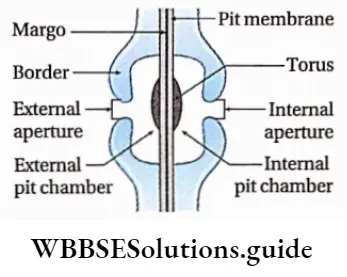
Unilateral compound pit
Sometimes two or more pits are found opposite to each other to form a large pit. It is called the unilateral compound pit. The cavity formed by breaking in the secondary wall is called the pit cavity.
Flower Anatomy
The primary cell wall and middle lamella that separate the two bordered pit cavities of a pit-pair are called the pit membrane or closing membrane. The pit opening is called the pit aperture. The empty region covered by the excess arching of the secondary wall is called the pit chamber.
The elevated over-arched secondary wall opens to the cell lumen by the pit aperture. As the secondary wall is usually very thick, a canal is formed in between the pit chamber and cell lumen called the pit canal.
The pit canal opens to the cell lumen and pit chamber by the inner aperture and outer aperture respectively. In front view the bordered pits exhibit two circles, the pit cavity forming a border around the pit aperture, and hence the name.
Vessels or tracheae: vessels or tracheae are tubular, thick-walled, non-living members of the xylem tissue.
Origin of vessels: Vessels of the primary xylem originate from the procambium and those of the secondary xylem develop from the cambium. The vessels evolved from long and narrow tracheids.
Vessels or tracheae Distribution: Vessels predominate in the vascular tissues of most of the angiosperms. Vessels are absent in Trochodendron, Tetracentron, Amborella, Takhtajania, etc., plants. They are absent in pteridophytes except in Selaginella, Equisetum, Pteridium.
They are also absent in most of the gymnosperms. Gnetum, a gymnosperm, contains vessels in its stem. They are present in both the primary and secondary xylem of angiosperms.
Handwritten notes on anatomy of flowering plants PDF
Vessels or tracheae Structure:
- The elongated, cylindrical vessels are dead at the matured stage.
- They are joined end to end and remain arranged in vertical rows.
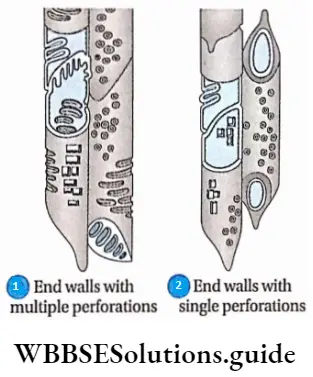
- The transverse partition walls or end walls dissolve at the matured stage and form a true tubular structure or tracheae.
- The end walls have a number of small holes at the surface. This type of end wall is called a perforation plate.
- The pattern of perforations may be of two types. These are—
- Simple, with a single large pore at the end (example Quercus sp.) and
- Multiple, with more than one pore, multiple perforations may be of three types
- Scalariform, with multiple pores arranged in a ladder-like manner (example Liriodendron sp.);
- Foraminal, with a number of pores arranged in a circular pattern (for example Ephedra sp.); and (reticulate, with a network of small pores (for example Rhoeo sp.).
- They also have numerous pits on their lateral walls.
- The vessel elements run parallel to the long axis of the plant parts in which they occur.
- Tracheae possess thick lignified cell walls.
- Vessels may be present as single or in groups. The groups may be arranged in radial, oblique, or tangential lines to the main axis of the plant.
Vessels or tracheae Functions:
- Their main function is the quick conduction of water and dissolved minerals.
- They also provide mechanical strength to the plants.
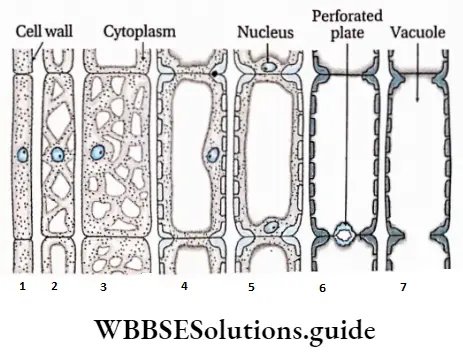
Different types of perforated plates in the trachea
Simple perforation plate: The perforation plate with a single large pore is known as a simple perforation plate.
Complex perforation plate: A perforation plate with more than one pore is known as complex perforation plate.
Scalariform perforation plate: The perforation plate with oval pores one above the other and separated by transverse bar of perforation plate is known as scalariform perforation plate.
Reticulate perforation plate: The perforation plate with pores arranged in a net-like or reticulate pattern is known as a reticulate perforation plate.
Xylem parenchyma: The parenchyma cells that occur as elements of the xylem tissue are termed xylem parenchyma or wood parenchyma.
Xylem parenchyma Origin: Xylem parenchyma originates from procambium. In the secondary xylem, the medullary ray parenchyma cells originate from the ray initials of the cambium.
Xylem parenchyma Distribution: Xylem parenchyma occurs in the primary and secondary xylem. These are found in all the gymnosperms and angiosperms.
Xylem parenchyma Structure:
- Xylem parenchyma cells may be oval, round, rectangular or square, elongated and sometimes irregular in shape usually with thin primary walls.
- Sometimes the wall becomes thick due to lignin deposition over the primary cell wall and forms simple pits.
- The pit pairs between the parenchyma and tracheary elements may be simple, half-bordered and bordered.
- Reserve foods in xylem parenchyma are mainly starch and fat. Crystals and tannins are also found in these cells.
- The presence of chlorophyll is also reported in some herbs and deciduous trees.
- The xylem parenchyma cells are oriented vertically or horizontally.
- Sometimes parenchyma cells protrude into vessels through pit cavities to form a balloon-like structure called tyloses.
Xylem parenchyma Function:
- It helps in the transport of water and minerals.
- It stores reserve food in the form of starch and fat and ergastic substances such as, oils, gums, resin, tannins, silica bodies, crystals, etc.
- The thick-walled lignified parenchyma also provides mechanical support to the plants.
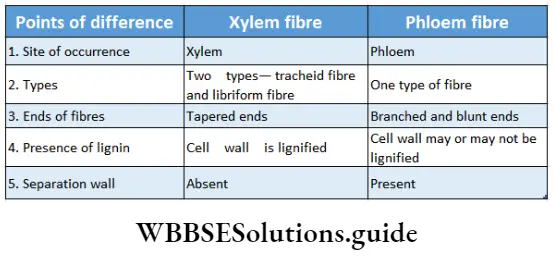
Xylem fibre: The dead sclerenchyma fibre associated with the xylem is known as xylem fibre or wood fibre.
Xylem fibre Origin: Fibres originate from the procambium in the case of the primary xylem whereas those of the secondary xylem develop from the fusiform initial of the cambium.
Xylem fibre Distribution: They are mostly found in vascular bundles of woody dicotyledonous plants. This type of fibre is present in primary and secondary xylem.
Xylem fibre Structure: Xylary fibres may be septate or aseptate. In tension wood, the xylem fibres are ofgelatinoustype.
Xylem fibre Types: Xylem fibres are of two types—libriform fibre and fibre tracheid.
- The libriform fibre is longer and thick-walled with simple pits.
- The fibre tracheids are smaller xylem fibres with bordered pits and are only found in the woody parts of the dicotyledons.
Xylem Fibre Function: The xylem fibres are responsible for mechanical support. They also store reserved food.
Xylem fibre Types of xylem: Based on origin xylem is of two types— primary and secondary xylem.
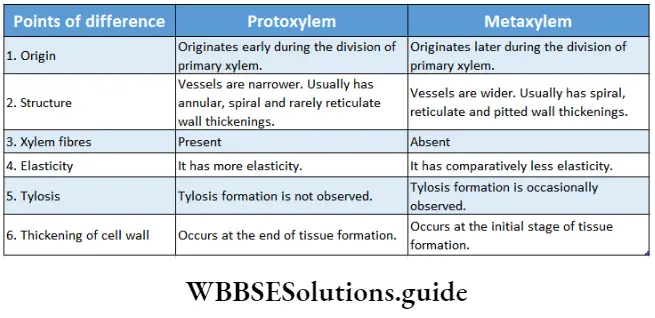
1. Primary xylem: The xylem that originates from procambium during the primary growth of the plants, is known as the primary xylem. On the basis of the structure and nature of the division, the primary xylem is divided into two types— protoxylem and metaxylem.
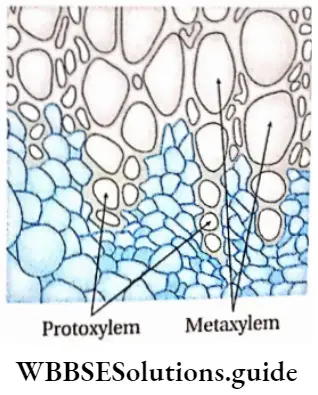
- Protoxylem is the xylem that forms first from the procambium and is known as protoxylem. The main components of this xylem are tracheids, trachea and xylem parenchyma. The protoxylem lacks xylem fibres. Tracheids and trachea consist narrow lumen.
- Metaxylem is the xylem, that forms later from the procambium, is known as metaxylem. The main components of this xylem are tracheids, trachea, xylem parenchyma and xylem ‘ fibres.
2. Secondary xylem: The xylem that originates from vascular cambium during secondary growth of the plants is known as secondary xylem. The secondary xylem is commonly known as wood.

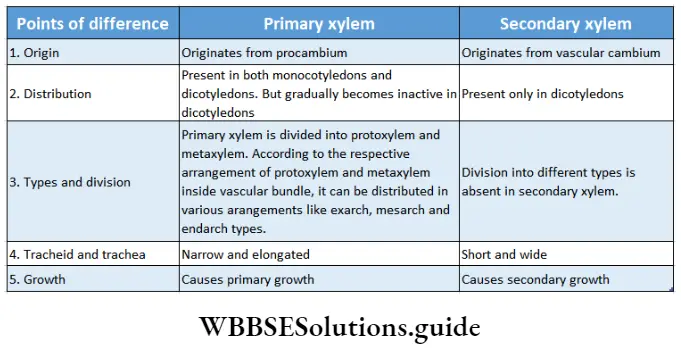
Stele and its types based on protoxylem and metaxylem arrangement
The stele is the central core of the plant axis containing the vascular and ground tissues and is delimited by the pericycle and endodermis respectively.
Four kinds of distributions are found in leaves, roots and stems on the basis of location of the protoxylem and metaxylem. They are—
Exarch: The xylem develops centripetally i.e., protoxylem remains towards the periphery and the metaxylem towards the centre.
Example: Root xylem.
Endarch: The xylem develops centrifugally, i.e., the protoxylem remains towards the centre and the metaxylem towards the periphery.
Example: Shoot xylem.
Mesarch: The xylem develops both centripetally and centrifugally i.e., the metaxylem is distributed on both regions (periphery and centre) and protoxylem is present between it.
Example: Leaf xylem.
Centrarch: Protoxylem is present at the centre and metaxylem surrounds it.
Example: Xylem of fern.

Phloem
Phloem Definition: Phloem is a complex, permanent tissue, found inside vascular bundles, through which food is transported from leaves to different parts of the plant.
Phloem Distribution: Phloem is a part of vascular bundles of root, stem and leaves of all vascular plants.
Phloem Function:
- Phloem primarily helps to transport food from leaves to other parts of the plant.
- Phloem may also add mechanical strength to the plant body.
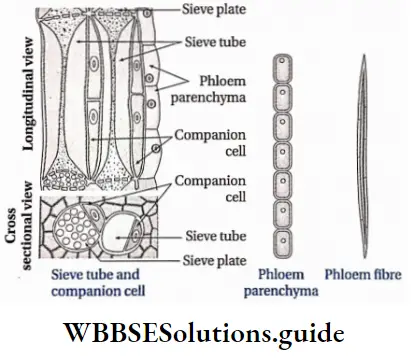
Phloem Components: Phloem is mainly composed of
- Sieve tubes or sieve cells,
- Companion cells
- Phloem parenchyma and
- Phloem fibres
Sclereids, laticiferous and resin ducts are also present in phloem tissues of some species. Phloem parenchyma, sieve tubes, companion cells and phloem fibres constitute the of phloem tissue in most of the dicotyledonous plants. Monocots plants do not have phloem parenchyma. In gymnosperm and pteridophytes, the phloem consists of sieve cells, phloem parenchyma and albuminous cells. The components are discussed below in separate heads.
The conducting components of the phloem are referred to as sieve elements that are characterised by the presence of sieve cells and sieve tubes.
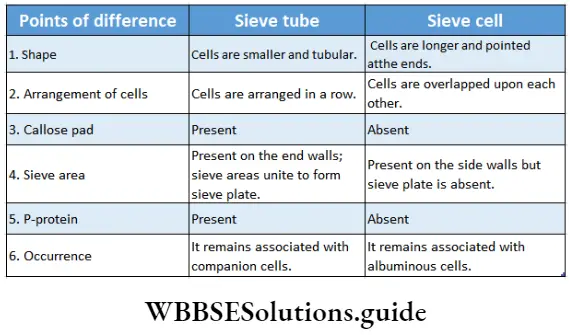
Sieve tube: The tube-like phloem cells containing sieve plates, which are the main food conducting phloem elements in angiosperms are known as sieve tubes.
Phloem Distribution: These are found in the secondary and primary phloem of angiosperms.
Phloem Structures:
- These are tube-like living cells, arranged longitudinally.
- The protoplasmic strand, present along the length of the cell is known as phloem protein or P-protein.
- The cell wall is thin and composed mainly of cellulose and pectin.
- The end walls have several perforations called sieve pores. An area with several sieve pores is called a sieve area.
- One or more sieve areas form sieve plates.
- The sieve tubes are non-nucleated. But in Smilax hispid, and Neptunia oleracea sieve tubes contain a nucleus.
- In winter deposition of polysaccharides, known as callose, covers the sieve pores.
- The thick layer of callose that blocks the sieve Pores is known as a callose Pad-
- Plastids occurring in the sieve tube protoplast may be either S-type or P-type depending on the nature of reserve food Starch accumulates in S-type whereas protein accumulates in P-type plastid.
Phloem Function:
- Helps in the conduction of food and important organic molecules like hormones etc., in angiosperms.
- Also helps in food storage.
Sieve cell: The elongated, living phloem cells with tapering ends are known as sieve cells.
Sieve cell Distribution: It is found in pteridophytes and gymnosperms
Sieve cell Structure:
- The sieve cells are arranged longitudinally.
- The cells are elongated and tapered at the ends.
- The cell wall is usually thin and made of cellulose.
- Sieve areas are present on lateral walls and sometimes on the end walls.
- A large central vacuole is present pushing the protoplast towards the wail forming the primordial utricle.
- Mitochondria, plastids and slimy proteinaceous structures or slime bodies are present.
- Starch grains are absent in sieve cells.
- They remain associated with albuminous cells instead of companion cells.
Companion cell: The elongated cells that have dense cytoplasm and remain associated with the sieve ted with the sieve tubes, are known as companion cells.
Companion cell Distribution: These are only found in angiosperms. Some parenchyma cells, similar to companion cells, that are associated with sieve tubes in ferns and gymnosperms are known as albuminous cells.
Companion cell Structure:
- These cells are associated with sieve tubes with the help of plasmodesmata. More than one companion cell can be associated with one sieve tube.
- The plasmodesmata connect the companion cells and the sieve tube through the primary pit field present between the two cells.
- They are usually shorter in length or may be as long as the associated sieve tubes
- The cells are vertically elongated.
- In some companion cells, wall materials deposit on the inner side of the primary wall to transform into transfer cell
- Prominent elongated or lobed nuclei are present in companion cells.
- The cells contain abundant Golgi apparatus, endoplasmic reticulum, mitochondria ribosomes, plastids, etc.
- In some companion cells P-proteins are found.
Companion cell Function:
- The companion cells are mainly related to the transportation of food through sieve tubes.
- These cells maintain the pressure gradient in the sieve tubes and help in lateral transportation. These cells serve as alternatives to sieve tubes.
Albuminous cell
The parenchyma cells, similar to companion cells, associated with the sieve cells in the gymnosperms are known as albuminous cells
Albuminous cell Structure:
- Albuminous cells are vertically elongated and may be of the same length as those of the sieve cells.
- Sieve and albuminous cells are connected through plasmodesmata
- Albuminous cells contain starch-free and protein-rich cytoplasm and occur at the margins of rays.
- Each of these cells contains a prominent nucleus and dense cytoplasm.
Albuminous cell Origin: In primary phloem, they develop either from procambium-derived phloem rays or from phloem parenchyma. In the secondary phloem, these cells originate from the vascular cambium
Albuminous cell Function: Helps in the conduction of proteins.
Phloem parenchyma: The parenchyma cells, other than albuminous and companion cells, found in phloem are called phloem parenchyma.
Phloem parenchyma Distribution: These are found in phloem tissues of dicotyledons, gymnosperms and pteridophytes. Phloem parenchyma is absent in monocots and a few members of Ranunculaceae.
Phloem parenchyma Structure:
- Phloem parenchyma cells are rectangular or rounded in the transverse section.
- In the longitudinal section, these cells appear oblong with rounded or tapered ends.
- The cell walls are thin and non-lignified with numerous pit fields.
- The cell wall is made up of cellulose.
- Sometimes scarified and thick-walled inactive parenchyma cells are observed.
- Phloem parenchyma cells with folded walls are known as transfer cells.
- These cells are the components of both primary and secondary phloem.
- In the primary phloem, the parenchyma cells remain parallel to the long axis of the associated xylem.
- In secondary phloem, they remain parallel or perpendicular to the long axis of the associated xylem.
Phloem parenchyma Function:
- Helps in organic food transport,
- Stores produced food and waste products.
Phloem fibre: The elongated sclerenchyma fibres associated with the phloem tissue are known as phloem fibres. The phloem fibres are the extrasolar fibres. They are also called bast fibres or bast wood fibres. These are the only dead elements in the phloem.
Phloem fibre Distribution: These are found in the primary and secondary phloem of angiosperms.
Phloem fibre Structure:
- These fibres are dead at maturity.
- They have a lignified, thick cell wall and are elongated with tapering ends, interlocked with each other. But in Linum sp. phloem fibre wall is not lignified.
- Fibre walls have simple pits with linear or round apertures. Sometimes, bordered pits are also found.
- The fibres may be septate or aseptate.
- In cross-section, they appear as isolated or scattered strands, as continuous or irregular bands, and as clusters over the phloem strand.
- They may form cylinders of tangential sheets encircling the inner tissues.
Phloem fibre Function:
- The phloem fibres give mechanical strength to the plants.
- They protect the inner tissues.
- Septate fibres may store starch, oils, resins, etc.
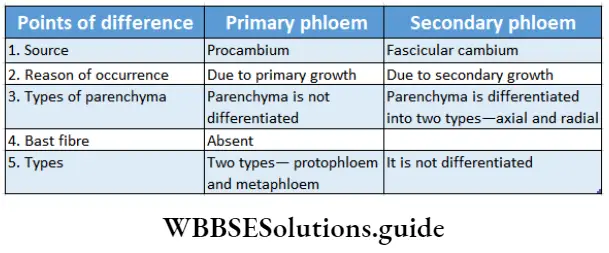
Phloem fibre Types of phloem: On the basis of origin, phloem is divided into primary phloem and secondary phloem.
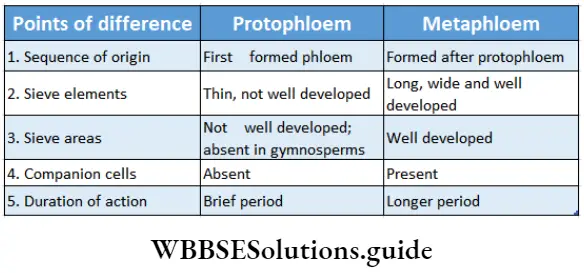
1. Primary phloem: The phloem that originates from apical procambium during primary growth is known as primary phloem. According to the sequence to development, the primary phloem is divided into protophloem and meta phloem.
- Protophloem is the phloem produced during the division and differentiation of procambium.
- Metaphloem is the phloem produced after the formation of protophloem during the division and differentiation of procambium.
2. Secondary phloem: The phloem that originates from fascicular cambium during secondary growth in mature plants, is known as secondary phloem.

Special tissue or Secretory tissue
Special tissue or Secretory tissue Definition: The special type of permanent tissue, composed of various types of cells that are present in clusters to carry out secretion or excretion in plants is called special tissue or secretory tissue.
Special tissue or Secretory tissue Characteristics:
- Transformed parenchymal cells cluster together to form secretory cells or glands.
- The rate of metabolism is high in cells of secretory tissue. This makes the protoplasm of parenchyma cells thick and granular.
- The glands either store the secreted substances in vacuoles or excrete them outside.
Special tissue or Secretory tissue Types: On the basis of position of occurrence, secretory tissues are of two types external glands and internal glands.
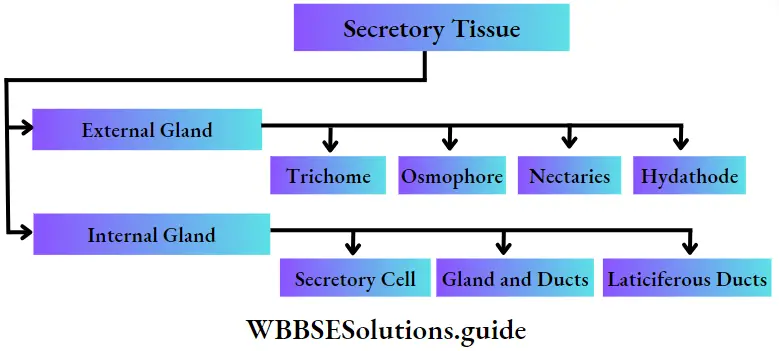
External glands: The glands or the secretory structures which are formed from the epidermis or hypodermis of plants and are present outside the plant body are called external glands.

External glands are of different types.
These are—
Trichome: These structures are present at the outermost layer or epidermis of the plant body. The root epidermis or epiblema does not have trichomes.
External glands Characteristics:
- Trichomes can be unicellular or multicellular,
- The wall of trichomes is thin,
- The apical cells of the trichome are involved in secretion.
External glands Function:
- These glands absorb metabolic substances,
- Mucilage and enzymes are secreted from them,
- They store water,
- They protect the plant from other animals. example Glandulartrichomesof tobacco plants, and trichomes in pitcher plants.
Nectaries: These structures are commonly associated with floral parts. But some extrafloral nectaries may also occur on vegetative parts such as different parts of flower, stem and leaves.
1. External glands Characteristics:
- The columnar cells of these structures are composed of dense cytoplasm and are rich in endoplasmic reticulum,
- The glands are multicellular and sessile (without stalk).
- Nectaries secrete a sugary fluid called nectar.
2. External glands Function:
- These structures secrete and store nectar,
- They attract insects for pollination with this nectar. example In dicot flowers, nectaries are present at the basal part of ovaries, stamens and perianths (sepals and petals). Nectaries are also found at the rim of the pitcher plant, on the leaves of Dolichos lablab.
Osmophore: These special glands are responsible for fragrance in various parts of the flower.
External glands Characteristics:
- The shape of glands of different types, like—tongue-shaped, brush-shaped, flap-shaped, or cilia etc.
- The glands are multicellular and have intercellular spaces.
- Volatile aromatic substances(olis) are secreted by these glands and may vaporise immediately or may remain as droplets.
External glands Function: The scent of the gland attracts insects which is helpful for pollination. example These structures are found on sepals and petals of species of Restrepia.
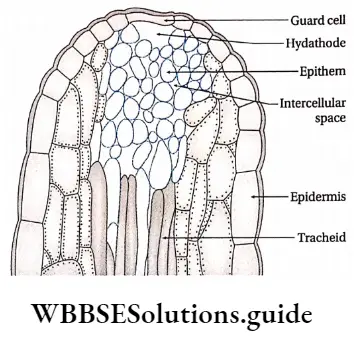
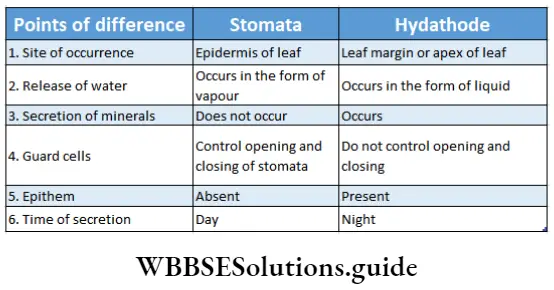
Hydathode: It is present in the serrated leaf margin of herbs where veins and venules get terminated.
External glands Characteristics:
- Hydathodes exudate water under conditions if low rate of transpiration and high root pressure.
- Each hydathode contains either one or more than one pore. A water cavity remains associated with each pore.
- Each of them contains a tissue of small, thin-walled, parenchymal cells with dense cytoplasm and profuse intercellular spaces. This tissue is called epithem.
- Epithem lacks chlorophyll and cells are associated with terminated ends of tracheids at vein-endings.
- Guard cells are present at the terminal end beneath which stomata are present. These are incapable of opening and closing.
- On both the sides of tracheids, chlorenchyma tissues are present.
External glands Function: Water and dissolved salts in it are forced out from tracheids and flow through epithem. Then this sap comes out through the stomata. At dawn, this water with soluble salts is exudated through the hydathode as dew.
Example: Hydathode is seen in tomato plants, grass, etc.
Internal glands: The secretory glands which are present within the different tissues of different parts of the plant are called internal glands.
Internal glands are of many types, like—
1. Secretory cells: The cells are different from adjacent cells as they contain a variety of substances.
2. Characteristics: These cells are larger in size, isodiametric or elongated into sacs or tubes.
Internal glands Function: Cells may contain balsams, resins, oils, gums, mucilages, crystals, etc.
- Needle-shaped crystals of calcium oxalate deposition are found in idioblast cells in the petiole of Colocasia. These crystals are known as raphides.
- In Ficus leaf calcium carbonate deposition is found in the specialised epidermal cells called lithocysts.
Glands and ducts: Secretory materials remain stored within the large, more or less isodiametric cavities’ or elongated canals. These cavities or canals are known as glands or ducts respectively.
Internal glands Characteristics:
- Some cells having thin cell walls and dense cytoplasm, associate together to take part in internal secretion.
- Secretory substances from the protoplast of the cell deposit in the inner cavity of the glands or ducts.
Internal glands Function:
- Glands are the source of various essential oils.
- Resin is deposited in resin ducts.
Examples: the Oil gland of Eucalyptus, oil gland of cotton seed, rubber, and resin duct of banyan.
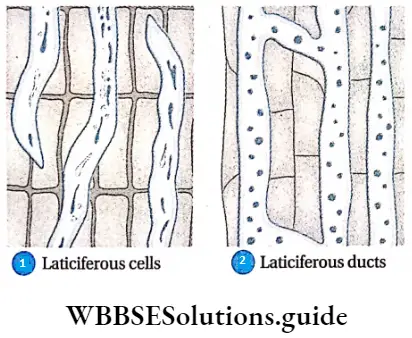
Laticiferous duct: The most important of all plant secretions is latex.
Internal glands Characteristics:
- The duct or tube-like structure, which secretes and stores latex, is called a laticiferous duct.
- This duct is thin-walled and has many nuclei.
- The unbranched, unicellular laticiferous duct is called a non-articulated laticiferous duct laticiferous cell or latex cell.
- When branched laticiferous cells sometimes form a network by partial or total dissolution of their end walls. These are called articulate laticiferous ducts or laticiferous vessels.
- Latex is a white or yellow, more or less viscous fluid. Latex contains emulsion of proteins, sugars, gums, alkaloids, enzymes, etc.
Internal glands Function:
- Latex in laticifers is mainly used as a protection measure against herbivorous animals.
- Latex is economically very important, as it is used to produce rubber. example, Laticifer cells are present in the stems of banyan trees. The laticiferous vessel is present in Hevea sp.(rubber plant), papaya, tobacco, etc.
Tissue Systems
Tissue Systems Definition: A system in which a single tissue or different tissues aggregate to perform specific functions, irrespective of their position in the plant body is known as a tissue system.
On the basis of location and function, Sachs (1875) proposed three types of tissue systems in higher plant’s body—
- Epidermal tissue system,
- Ground or fundamental tissue system and
- Vasculartissue system.
Epidermal tissue system
Epidermal tissue system Definition: The epidermal tissue system is the outermost continuous layer or layers of cells of all the plant parts that protect the inner tissues.
Epidermal Tissue System Origin:
- The epidermal tissue system is developed from the protoderm of the apical meristem.
- Epiblema is formed from the layer of apical meristem of the root, covered with root cap.
- These tissues are formed by the anticlinal division of the cells present in the protoderm.
Epidermal tissue system Structure: The components of the epidermal tissue system are of different nature. The main structural components are—epidermis, epidermal outgrowths and epidermal openings.
Epidermal cells
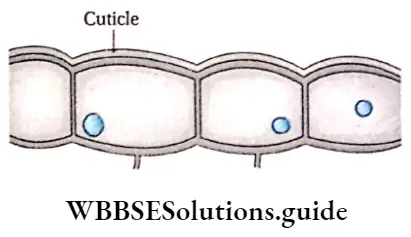
Epidermal cells Definition: The external protective cell layer of the whole plant body except the roots, which stays in direct contact with the environment is known as the epidermis.
Epidermal cells Characteristics:
- The epidermis is formed of living parenchyma cells.
- Cells are tubular or oval, closely compact without intercellular spaces.
- The cell wall is composed of cellulose.
- The epidermis is composed of a single layer of cells.
- In some plants epidermis is multi-layered, known as multiple epidermis. This type of epidermis is observed in the leaves of Ficus sp. and Nerium sp.
- The multiple epidermis is formed through the periclinal division of the epidermal initials.
- The multiple epidermis of orchid roots is known as velamen.
- Epidermal cell walls are usually thin. Thick-walled lignified epidermal cells occur in some gymnosperms.
- The cuticle layer is formed on the outer surface of the leaf and stems by the deposition of cutin.
- Sometimes wax may be deposited on the surface of the cuticle.
- Generally, chloroplast is absent in the epidermal cells, but in ferns epidermal cells and guard cells of stomata. Bulliform contain chloroplast.
- Mostly the epidermis is continuous but the epidermis of leaves is discontinued by the stomata, whereas, in some stems, it is broken due to the presence of lenticels.
Epidermal Cell Types: Different types are discussed below.
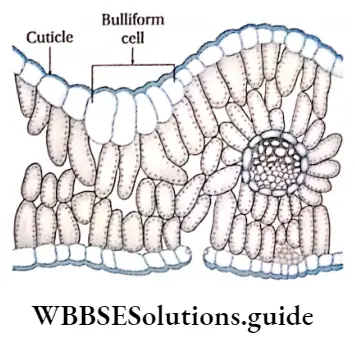
1. Bulliform cells or motor cells: These are large epidermal cells found in the upper epidermis of the leaves in monocotyledons. These cells are bubble-shaped and occur in groups. The outermost wall of these cells is cutinised and covered with cuticle. The bulliform cells provide support to the leaves during development. Bulliform cells cause the unrolling of developing leaves and movement of mature leaves by developing rhythmic turgor pressure. These cells serve as water reservoirs.
2. Silica and cork cells: Two types of epidermal, short cells that remain associated with long epidermal cells, are found in grasses. Some cells are filled with silica and are known as silica cells. The rest of the cells contain solid organic substances and are known as cork cells. Cork cells have suberised walls.
3. Myrosin cells: The elongated, sac-like cells found in some dicot plants. These cells are secretory in nature and contain myrosin enzymes. They are also known as myrosin cells.
4. Lithocysts: The leaf epidermal cells of certain plants (for example Ficus) contain crystals of calcium carbonate (known as cystoliths). These cells are called lithocysts.
5. Sclereids: The epidermal cells of seed coat in some leguminous plants and scale epidermis of garlic are composed of sclereids.

Epidermal cells Function:
- It protects the inner tissues from any adverse external factors like high temperature, desiccation, mechanical injury, pathogenic infections, etc.
- The cuticle protects plants from desiccation, as it is impervious to water.
- Wax is deposited on the inner surface of the pitcher of Nepenthes sp. (pitcher plant) in the form of overlapping scales, where insects stick easily.
- Serves as water storage and acts as secretory cells.
- Chlorophyllous epidermal cells are involved in photosynthesis.
Epidermal outgrowth
Epidermal outgrowth Definition: Various protuberances that arise from the epidermal cells in plants irrespective of their structures and functions are known as epidermal outgrowth.
Epidermal outgrowths are present in roots, stems, leaves, floral parts, seeds and stamens. Collectively these outgrowths are known as trichomes.
Epidermal outgrowth Types: These are the unicellular or multicellular, hairy or glandular, simple or branched outgrowths of epidermal cells. The glandular ones are secretory in function. The hairy trichomes provide protection and also prevent water loss. The trichomes can be stellate (star-like) or dendroid (a miniature tree form, for example, Verbascum).
They may also occur in tufts (for example Hamamelis). Hairy projections are found in leaves, roots, stems and also in flower petals. The hairy outgrowth of flower petals is known as papillae. Different types of epidermal outgrowth are discussed below.
1. Stem hair: These are unicellular or multicellular hairy outgrowths present on the stem. They protect the plants from various unfavourable conditions.
2. Root hair: Root hairs are always unicellular and are present just behind the root tip of most monocots and dicots. They arise from distinct epidermal cells termed as trichoblasts, which protrude out of the root surface to form unicellular root hairs. The main function of root hairs is to absorb water and minerals in addition to anchoring the plant to the soil.
Glandular trichomes: These are the multicellular glandular outgrowths of epidermal cells. They are specially found in the digestive glands of insectivorous plants like Drosera sp.

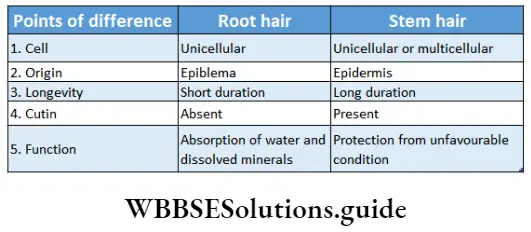
4. Stinging hair: These types of outgrowth are special kinds of hairs which are provided with a bladder-like broad base and capillary tube-like apical cell. The apical cells of these hairs become elongated and filled with poisonous juice. If any animal touches these hairs, it will come in contact with the poisonous juice, which causes irritation. These hairs are found in plants such as Mucuna sp., Tragia sp., etc.
5. Water vesicles: These are swollen epidermal cells which form a bladder-like structure. These vesicles serve as water storage organs. These are found in the ‘ice plant’ (Mesembryanthemum crystallinum).
6. Scale or peltate hair: These are multicellular, flat, non-glandular hairs with or without stalk (i.e., sessile). These are formed of disc-like cell plates. The stalked hairs are known as peltate hair and the hairs without stalk or sessile are known as scale hair.
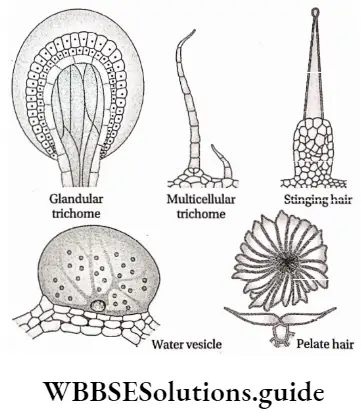
Epidermal Outgrowth Function:
- Root hairs and stem hairs protect different parts of the plants.
- The non-glandular hairs of the stem and leaves reduce the rate of transpiration.
- Glandular hairs protect the plants from different herbivorous animals.
- Epidermal hairs help in pollination.
- Hairs found on the fruits and seeds help in their dispersal.
- Root hairs help in the conduction of water and minerals from the soil.
- Sometimes epidermal hairs store water.
Epidermal Openings: The Epidermis of aerial parts of plants, mainly in leaves, is not continuous at all. It is interrupted by various openings. Different types of openings found in leaves are discussed below.
Stomata: Stomata (singular: stoma) are the microscopic pores found on the epidermal surface of aerial parts in higher plants. The term stoma was first coined by de Candolle in 1827, which means mouth.
Epidermal Openings Distribution:
- Stomata occur abundantly throughout the surface of the lamina except the vein areas. When the lamina is very thick, stomata may also occur along the veins.
- Stomata are present either in the lower or upper epidermis or on both the epidermises of leaves.
- If stomata are present on both surfaces of a leaf with fewer stomata on the upper surface, then the leaf is called an amphistomatous leaf.
- Leaves with stomata only on the lower surface are called hypostomatous leaves. Floating leaves and partially submerged leaves contain stomata only on the upper surface of the leaves. This type of leaf is called epistomatous leaf.
- The stomata may be located in pits (Ammonophilia arenaria), below the epidermal leaf surface example Pinus). In xerophytes, such as Nerium, Xanthorrhoea, etc., stomata are present in the subepidermal cavity. These types of stomata are known as sunken stomata. Sometimes these cavities are lined with trichomes.
- Stomata are present on the epidermis of the calyx, corolla, androecium and gynoecium.
- In Saxifraga stolonifera stomata are located on raised patches and project above the level of the leaf surface. This type of stomata is called raised stomata.
NEET biology anatomy of flowering plants revision notes
Epidermal Openings Structure:
- A stoma consists of a small stomatal aperture(pore) and two guard cells.
- Each pore is bounded by two specialised kidney-shaped or semilunar epidermal cells called guard cells,
- The guard cells are surrounded by a certain number of epidermal cells known as the subsidiary or accessory cells,
- They together constitute the stomatal complex or stomatal apparatus,
- The stomatal aperture opens below into a large cavity, known as the stomatal cavity or substomatal chamber. This chamber remains in connection with the internal intercellular space system,
- Each stoma has prominent four sides. The thick ventral side faces the pore, the thin dorsal side towards the subsidiary cell, the upper lateral side faces the atmosphere and the lower lateral side faces the stomatal cavity,
- The cellulose microfibrils orient themselves radially in a semilunar guard cell wall called radial micellation.

Epidermal openings Function:
- The exchange of gases in plants occurs through stomata.
- Transpiration also occurs through stomata.
- Stomata also help in photosynthesis as the guard cell contains chloroplasts.
Hydathode or water stomata: Some specialised cells are present in the leaves of certain plants, that help to remove water and salts (dissolved in water) from the plant’s cells. They are known as hydathodes.
Ground or Fundamental Tissue System
The ground tissue system is the largest tissue system in the plant body. Ground tissue system is heterogeneous in nature, including diverse types of tissues specialised for different functions.
Fundamental Tissue System Definition: The tissue system including all the tissues of the plant body, except the epidermal and vascular tissues, is known as the ground or fundamental tissue system.
Fundamental Tissue System Origin: All the tissues of the ground tissue system develop from the ground meristem of the embryo.
Fundamental Tissue System Division:
The ground or fundamental tissue system is divided into two regions—
- Extrastellar region or extra stellar ground meristem and
- Intrastellar region or intrastellar ground meristem.
The group of tissues present between the pith and pericycle is known as stele. The ground tissue outside the stele is known as extrasolar ground tissue and that inside the stele is known as interstellar ground tissue. Both the tissue zones are further differentiated for certain particular functions. In leaves, the ground tissue present in between the upper and lower epidermis is known as mesophyll tissue.
Estrasteilar region: This region is formed of four parts. They are—
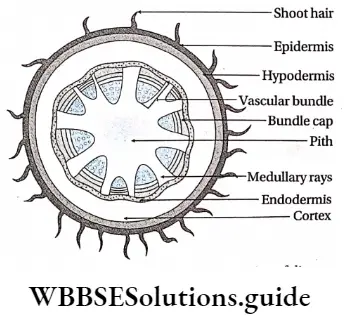
Hypodermis: The 2-3 cell layered thick tissues found just below the stem epidermis is known as hypodermis.
1. Estrasteilar region Characteristics:
- In stems of dicotyledons, the hypodermis is formed of collenchyma cells and in stems of monocotyledons, it is formed of sclerenchyma cells,
- Hypodermis is absent in roots and leaves.
2. Estrasteilar region Functions:
- Hypodermis provides mechanical support to the stem,
- It protects the internal tissues of the stem,
- It helps in gaseous exchange between the environment and the cortex.
- Chloroplast containing collenchyma cells help in photosynthesis.
Cortex: The region in between the epidermis (in case of monocot stem) or hypodermis (in case of dicot stem) or epiblema (in case of monocot and dicot roots) and endodermis, is called cortex. This is composed of many layers of thin-walled parenchymatous cells. Sometimes cortex in the stem of dicotyledonous plants contains collenchymatous cells.
1. Estrasteilar region Characteristics:
- The cortex, in the stem, is composed of parenchyma and/or collenchyma cells. Hence it can be heterogeneous in nature.
- In roots, the cortex is formed of only parenchyma cells, hence it is homogeneous in nature.
- In dicot stems, it is present between the hypodermis and the starch sheath. In the monocot stem, it is present between the epidermis and endodermis.
- Cortex is present between the epiblema and endodermis in both monocot and dicot roots.
- The parenchyma cells of the stem and root cortex contain colourless leucoplastids, but the parenchyma cells of the leaf cortex bear chloroplastids.
- In some cases, the stem cortex contains resin ducts.
2. Estrasteilar region Functions:
- It acts as a water and food storage.
- It helps in the conduction of water by maintaining the water potential.
- Sometimes it also acts as photosynthetic tissue due to the presence of ch|oroplastids.
Endodermis: The innermost layer of the cortex or the outermost layer of stele is the endodermis. It is the separation of the cortex from the stele. It is composed of a single layer of barrel-shaped parenchymatous cells.
Estrasteilar region Characteristics:
- The endodermis of many dicotyledonous stems may contain starch granules. Such endodermis is known as a starch sheath.
- Endodermis is prominent in underground stems.
- In roots, the endodermal cells are thick-walled. Lignin, suberin, and cutin present in the cells form strip-like structures near the cell wall which are known as Casparian strips.
- It usually surrounds the entire stele.
- In polysialic (more than one stele) conditions it surrounds the vascular tissue of each stele individually (for example Nymphaea)
Estrasteilar region Functions:
- It protects the interstellar region.
- Sometimes, endodermis stores starch, protein granules, fats and tannins.
- The thick cell wall of the endodermis serves as a barrier for heavy metal transport into or out of vascular tissues.
- In rhizomes, it controls water transport between the stele and the cortex.
- Endodermal cells accumulate various metabolic substances like benzoquinones, naphthoquinones, anthraquinones, etc. These are called secondary metabolites. These have anti-pathogenic activity. Thus, endodermis protects the interstellar zone from different pathogens by forming a barrier.
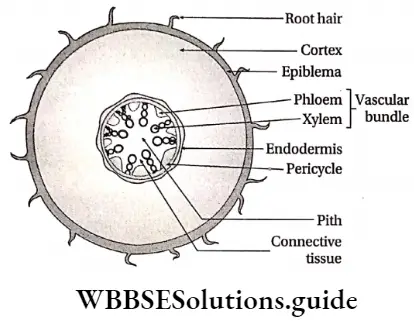
Mesophyll: The region between the upper and lower epidermis in leaves, formed of chloroplast containing parenchymatous cells, is known as mesophyll.
Mesophyll Characteristics:
- In dicotyledonous leaves, this region is made of palisade and spongy parenchymatous cells. In monocotyledonous leaves, this region is made of spongy parenchymatous cells only. These cells are chlorenchymatous.
- Palisade parenchyma cells form two or three layers near the upper epidermis. These cells are longer than their breadth with rounded ends. They are closely packed.
- Spongy parenchyma cells are oval or spherical and are loosely arranged.
Mesophyll Functions:
- These tissues mainly produce and store food,
- They play an important role in gaseous exchange and transpiration.
Interstellar region: This region is formed mainly of three parts.
Pericycle: The pericycle is the region immediately inner to the endodermis, surrounding the vascular tissues.
Interstellar region Characteristics:
- It is formed of one or more layers of cells,
- The pericycle typically consists of parenchymatous cells as found in the roots of all vascular plants and stems of pteridophytes.
- In dicotyledonous stems, the pericycle is multilayered and formed of sclerenchyma. These sclerenchyma cells form the bundle cap above the vascular bundle.
- Sometimes discrete bands of sclerenchyma fibres, are also present in pericycle.
- Pericycle gives rise to lateral roots.
- Pericycle is absent in the stems and roots of aquatic plants.
Interstellar region Functions:
- In roots, the pericycle gives rise to adventitious and lateral roots.
- Pericycle of stems store food! and provide mechanical support to the plants.
- Secondary meristematic tissues are formed from the pericycle.
Pith: Pith is parenchymatous ground tissue located at the centre of the stem or root axis. The pith is also called the medulla.
Interstellar region Characteristics:
- It is generally parenchymatous with profuse intercellular spaces. In certain monocots (for example Canna) pith is sclerenchymatous.
- The pith cells are usually isodiametric and sometimes remain arranged in longitudinal series,
- The thin-walled cells usually contain colourless leucoplasts.
- The outer pith cells are smaller with thicker walls containing dense cytoplasm. They form a distinct zone perimedullary zone or medullary sheath.
- Pith is very thin and inconspicuous in dicot root. Pith is even absent in many dicotyledonous roots.
- In the plants of the family Cucurbitaceae and many grasses, hollow piths may be formed with broken wall lining.
Interstellar region Functions:
- Sclerenchymatous pith provides mechanical strength to the plants,
- The pith cells may store starch, fatty substances, crystals and tannins.
Medullary rays and conjunctive tissue: Extension of pith in the form of narrow parenchymatous strips, present in the interfascicular regions i.e. in between the vascular bundles, called the medullary rays.
The parenchyma cells, present around the xylem and phloem in the root, form conjunctive tissue.
1. Interstellar region Characteristics:
- Each cell contains dense protoplasm, a well-developed nucleus and different other components,
- Medullary rays connect the pith and cortex.
2. Interstellar region Functions:
- Interstellar secondary growth occurs through the formation of interfascicular cambium.
- It helps in the transportation of dissolved substances.
- It also stores water and food.
Vascular Tissue System
Vascular Tissue System Definition: The tissue system, that consists of the xylem and phloem and helps in the transportation of water and food in plants, is known as the vascular tissue system.
The xylem and phloem constitute discrete conducting strands called vascular bundles. Each bundle is the isolated unit of conducting tissues consisting of xylem and phloem, covered frequently with a sheath of thick-walled cells.
In spite of the mechanical support the vascular bundles primarily function in conduction, xylem for the conduction of water with dissolved mineral matters, and a phloem for the conduction of prepared food matters in solution.
Vascular Tissue System Origin: Vascular bundles originate from the primary meristem. The vascular bundle is formed of the primary xylem and phloem. The primary xylem and phloem originate from procambium.
In dicotyledons, the vascular bundle is arranged radially. In monocotyledons, the vascular bundles are arranged non-uniformly with the ground meristem.
In the case of roots, the xylem and phloem are arranged in different radii to form a stele. In stems either they stay scattered throughout the ground tissue or may stay arranged in a compact ring to form the stele.
In some abnormal cases, vascular bundles may also be formed in the cortex or pith to form cortical and medullary bundles respectively.
Vascular Tissue System Structural components of vascular bundles:
The main three components of vascular bundles are— primary xylem, primary phloem and cambium.
- Primary xylem: The xylem formed during the primary growth of plants is known as primary xylem. The initially formed xylem with narrow lumen is known as protoxylem and the xylem formed later with wide lumen is known as metaxylem.
- Primary phloem: The phloem which occurs during the primary growth of plants is known as primary phloem. The initially formed phloem is known as. protophloem and the phloem that is formed later is known as metaphloem.
- Cambium: It is the region between the xylem and phloem, present in the dicotyledons and angiosperms. Cambium is not present in monocotyledons.
Types of vascular bundle: According to the arrangement of the xylem and phloem tissues,
The following types of vascular bundles are found—
Conjoint vascular bundle: In conjoint type, the xylem I and phloem lie together in the same radius. This type of vascular bundle is of three types.
They are as follows—
Collateral vascular bundle: In the collateral vascular bundle, the xylem and phloem tissues remain side by side on the same radius with the phloem being external to the xylem.
Again depending on the presence or absence of the Cambium
The vascular bundles are of the following types—
- Closed collateral bundle, the cambium is absent in between the xylem and phloem in this type of vascular system. Stems with this type of bundle do not show normal secondary growth. These closed vascular bundles usually remain enclosed in the sclerenchymatous bundle sheath. Generally, this type of vascular bundle is found in monocotyledonous stems.
- Open collateral bundle, here the cambium is present in between the xylem and phloem tissues. This cambium is called fascicular cambium. This type of vascular bundle is found in dicotyledonous stem.
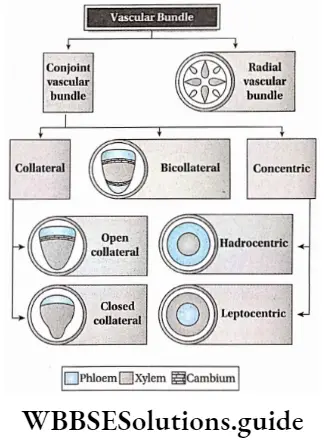
Bicollateral bundle: In this type of vascular tissue, phloem tissue is situated on both the peripheral and central regions of the xylem tissue. The phloem tissues are separated from the central xylem tissue by two strips of cambium (outer and inner cambium).
Plant Tissue And Anatomy Class 11 Notes
So the sequence of vascular tissues in bicollateral bundles from the periphery is outer phloem, outer cambium, xylem, inner cambium and inner phloem. These bundles are open type as two cambium strips are present.
Concentric bundle: In this type of vascular bundle, one of the vascular tissues completely surrounds the other. The concentric bundles are closed, as there is no cambium, in between the xylem and phloem tissues.
This type of vascular bundle is of the following types—
- Bundle, in this type, the xylem surrounds the central strand of the phloem. This type of vascular bundle is also termed as leptocentric vascular bundle. This type of bundle is found in the stems of Dracaena, Yucca, etc.
- Amphicribral bundle, in this. type the phloem surrounds the central strand of the xylem. Ills also known as hadrocentric bundle. This type of vascular bundle is found in stems of Pteridophytes such as in Selaginella.
Radial vascular bundle: In this type of vascular bundle, the primary xylem and phloem strands remain separated from each other by non-vascular tissues. These two tissues are situated on ultimate radii- These types of bundles are characteristic of roots.
The function of the vascular tissue system:
- Conduction of water and dissolved minerals from the soil to different parts of the plants is the main function of the vascular tissue system.
- The xylem helps in the conduction of water |s.
- phloem helps to transport the food prepared in leaves, to other plant parts.
- Cambium helps in secondary growth in plants.
- The vascular bundle also provides mechanical support to the plants.
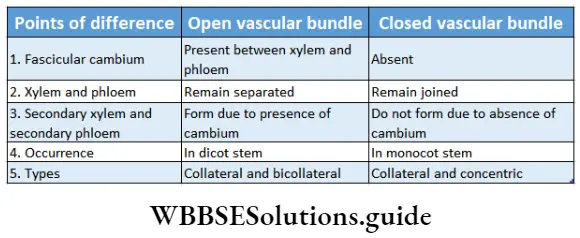
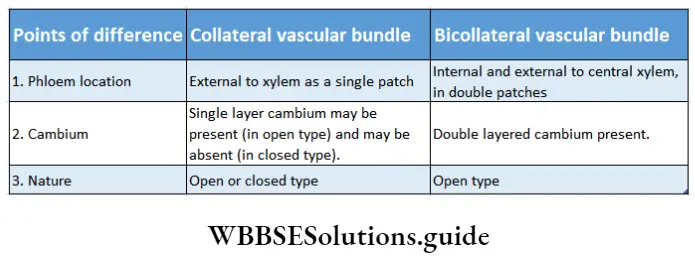

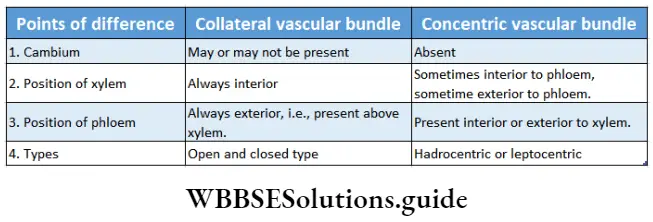
Secondary Growth In Plants
Secondary Growth In Plants Definition: After the primary growth, the increase in girth or thickness of plant parts due to the formation of secondary tissues by the activities of vascular cambium and phellogen, is known as secondary growth.
After the completion of primary growth, an increase in thickness is noticed in the woody gymnosperms, dicotyledons and monocotyledons. This increase in thickness occurs due to the formation of some new tissues by the activities of the lateral meristems such as cambium and phellogen (cork cambium).
The derived tissues are known as secondary tissues and the increase in girth or thickness of the plant parts is referred to as secondary growth.
The secondary tissues involved in the process are the secondary vascular tissues and periderm deriving their origin from the lateral meristems, cambium and phellogen or cork cambium respectively.
Cambium
Cambium Definition: The lateral secondary meristem, which helps in secondary growth in plants by the formation of secondary vascular tissues, is known as cambium.
Cambium Characteristics:
- Cambium is composed of thin-walled, protoplasm-filled cells that bear a well-developed nucleus.
- Cells divide parallel to the plant axis.
- Cambium is formed of two types of cells—
- Fusiform initials and
- Ray initials.
- Fusiform initials consist of tapering ends and ray initials are smaller in size.
Cambium Types: Different types of cambiums are described below.
Vascular cambium or Fascicular cambium
Vascular cambium Definition: The cambium, that is present at the vascular region of a plant to give rise to new vascular tissue, is known as vascular cambium.
Vascular cambium Types:
Vascular cambium is of two types—
1. Intrafascicular cambium: The cambium, present between the xylem and phloem of the vascular bundle or fascicle of both dicot and monocot plants, is known as intrafascicular or fascicular cambium. This cambium divides to form a secondary xylem in the inner side of the vascular bundle and a secondary phloem on the outer side.
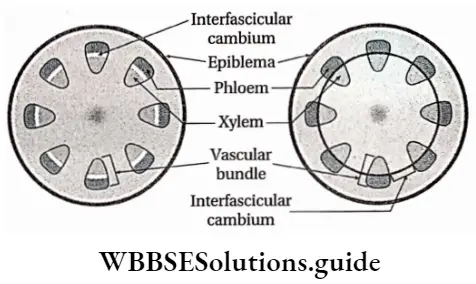
2. Interfascicular cambium: The cambium present in between two vascular bundles, is known as interfascicular cambium. During the secondary growth of the plants, the cells of the interfascicular cambium divide and combine with the intrafascicular cambium to form a cambium ring.
Cork cambium or Phellogen
Cork cambium Definition: When the parenchymatous cells of the cortex of the extra stellar region change to secondary meristem, then they are termed cork cambium or phellogen.
Cork cambium Function: Divides continuously to form phellem or cork at the outer side and phelloderm, at the inner side These two layers along with phellogen form the bark.
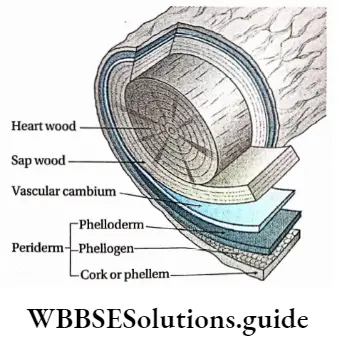
Secondary growth in typical dicotyledonous stems
In dicotyledons,’ secondary growth occurs in two regions
- Intrastellar region and
- Extrastellar region.
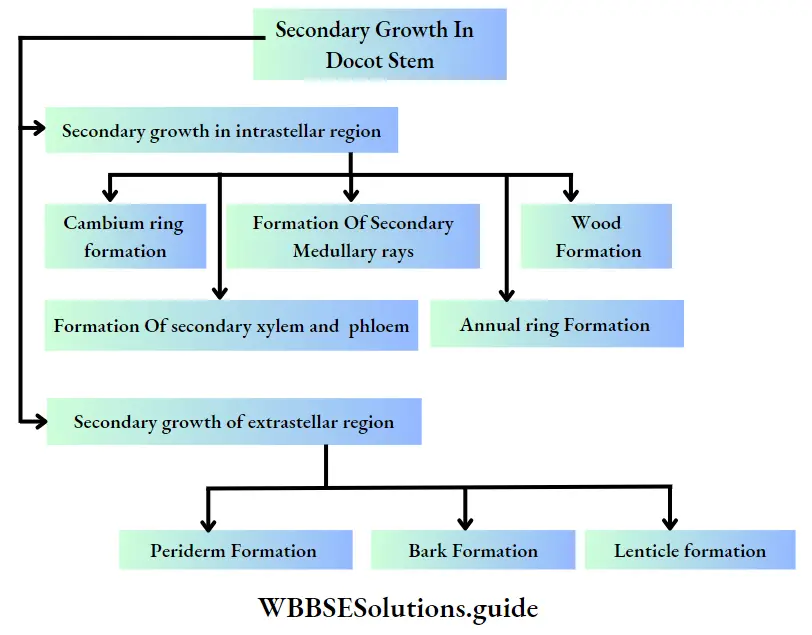
Secondary growth in interstellar region: In dicotyledons, secondary growth initiates in the interstellar region. The phases of growth are described below.
Formation of cambial ring:
- The fascicular cambium is present in the open vascular bundle of the stem.
- Each vascular is present in a discontinued stripe-like structure.
- The cells of medullary rays, present in the same plane as the fascicular cambium, divide and form strips of interfascicular cambium.
- The intrafascicular and interfascicular cambia unite to form a complete cambial ring.
Formation of secondary xylem and secondary phloem:
- The cambium ring divides and produces new cells in both of its inner and outer regions.
- The cells formed in the outer region transform into components of phloem and give rise to secondary phloem.
- The newly formed cells in the inner region of the cambium ring transform into xylem components and give rise to secondary xylem.
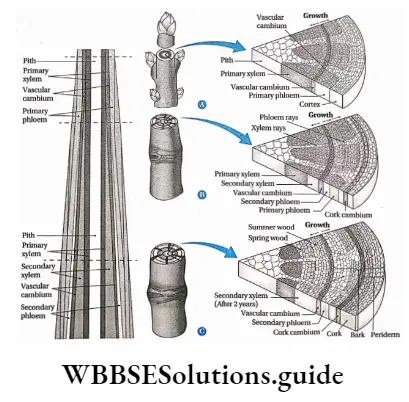
Formation of secondary medullary rays:
- Interfascicular cambium and fascicular cambium continuously divide to form a secondary xylem at the inner side and a secondary phloem at the outer side. Hence, medullary rays between the pith and cortex gradually decrease.
- In this condition, some cells of the interfascicular cambium produce strips of parenchyma cells radially and give rise to secondary medullary rays.
- These rays contain xylem rays and phloem rays.
Formation of annual ring:
- The activity of the cambium changes with seasons.
- In spring, cambium becomes more active which results in the production of a large amount of secondary xylem, i.e., wood.
- The wood formed during spring is less dense and is made up of vessels with wide diameters. This wood is called early wood.
- The activity of cambium gradually decreases in summer hence less amount of dense wood is produced. This part is made of a compact lignified xylem with narrow lumen. This wood is called latewood.
- In transverse views, these growth layers of cambium appear as rings and hence are referred to as growth rings. So, each growth ring represents one year’s growth. Hence, they are also known as annual rings. The age of a plant can be calculated by counting these concentric annual rings.

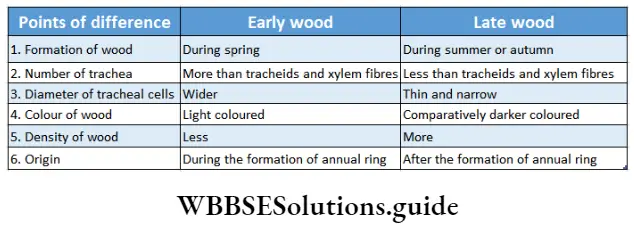
Types of wood: Wood is a hard, fibrous tissue found in the stems and roots of trees and other woody plants. Wood is sometimes defined as only the secondary xylem. In a living tree, it gives support to the plant.
Different types of wood are as follows—
Ring porous wood and diffuse-porous wood: Usually, large vessels occur in the early wood, making it more prominent than the latewood. The largest vessels exhibit a ring-like arrangement in the transverse section, this type of wood is called ring porous wood.
But in some plants, the vessels are found to be of more or less equal diameters. They remain uniformly distributed throughout the wood or throughout the growth ring during the gradual change from early to latewood. This type of wood is called diffuse-porous wood.

Sapwood and heartwood: After the formation of a considerable quantity of secondary xylem, two different types of wood zones appear in the stem. This wood zone is of two types—sapwood (or alburnum) and heartwood.
The outer region consisting of the recently formed xylem is called sapwood and the centrally located region which is formed earlier, is called heartwood. The colour of sapwood is lighter than the heartwood. Gradually, the sapwood gets transformed into heartwood.
Xylem parenchyma is present as a component of sapwood. the sapwood helps in the upward movement of water and nutrients, when sapwood transforms to heartwood, tyloses emerge from cells of sapwood.
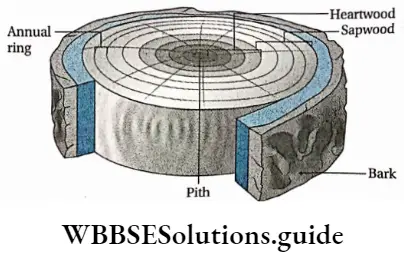
Softwood and hardwood:
- The secondary xylem of gymnosperms possesses 90-95% tracheid and rest 5-10% parenchyma cells. However, it lacks trachea and fibres. This type of wood is called softwood.
- The secondary xylem of angiosperms(dicots) is composed of trachea, xylem fibres and xylem parenchyma. It consists of 5-10% tracheid. This type of wood is called hardwood.
Tylosis
The secondary growth that occurs in plants for many years creates pressure on the earlier-formed xylem components (tracheid and trachea). The xylem parenchyma and ray parenchyma cells protrude like a balloon inside the tracheary element through the pit membrane of the half-bordered pits connecting the parenchyma and trachea.
These protrusions are known as tyloses (plural of tylosis). Tyloses get enlarged and may block the lumen of the tracheary elements. The nucleus and a small part of the cytoplasm enter into the tylosis.
Starch, resin and other substances may deposited in these balloons. In the beginning, the wall of the tyloses remains thin but later, it gets lignified.
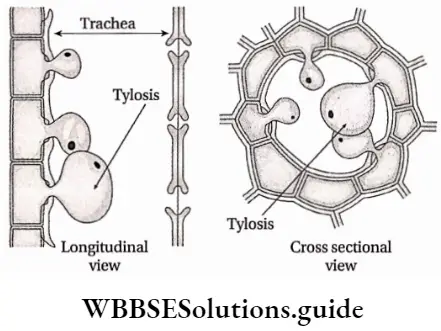
Secondary Growth in the Extrastellar Region
Formation of Periderm:
- In the interstellar region, due to the ceaseless formation of secondary tissues from the cambium cylinder, considerable pressure is exerted on the epidermis and also on other extra stellar tissues.
- The epidermis becomes stretched and often ruptures.
- Cork cambium or phellogen originates in the extrastellar region as a secondary meristem to withstand the above-mentioned pressure and to protect the internal parts which get exposed due to the ruptured epidermis.
- The cork cambium cells further divide and produce phellem on the outer side and phelloderm on the inner side.
- The three newly formed tissues— phellogen or cork cambium, phellem and phelloderm together form the periderm.
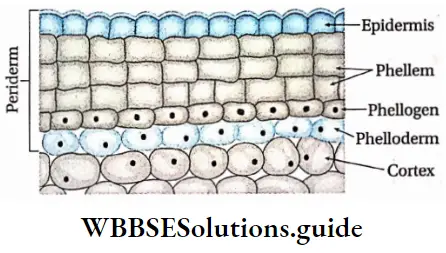
Formation of Bark:
- Additional layers of periderm are formed in the internal regions to withstand internal pressure.
- The cells of the outer layer move far away from the vascular bundles and they do not get sufficient amount of water and food.
- As a result, these cells of the outer periderm become dead on maturity. This dead outer layer of cells forms the bark of the tree.
- The cell walls of the outer cells of periderm have high suberin content, So they control the rate of transpiration.
- Bark protects the plants from heat, cold, pathogens and other stresses.
- Generally, successive layers of periderm are formed in the deeper regions as concentric rings surrounding the entire stem. This type of bark is known as ring bark.
- In some plants, the periderm is formed as overlapping scale-like layers.
- As a result, the outer tissues break up and are sloughed in patches. This is known as scale bark.
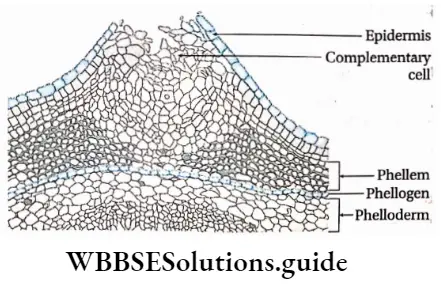
Formation of Lenticels:
- The periderm replaces the epidermis in respect to provide protection during a secondary increase in the thickness of the plant.
- The suberised wall of dead cork cells is partly impervious to gases. Thus, gaseous exchange between the internal living cells and the outer atmosphere becomes difficult.
- Some lens-shaped pores form on the surface of the stem and help in gaseous exchange. These pores are known as lenticels.
- These are formed with loosely arranged parenchymatous cells in the sub-stomatal region and those formed by phellogen, together are termed complementary cells.
- Its other components are phellem, phellogen and phelloderm.
- Only a few plants, mostly climbers, do not have lenticels though periderm is formed.
- Lenticels start forming just below the stomatal complex during primary growth preceding periderm formation. Lenticels protrude above the surrounding periderm due to their bigger size and loose arrangement of cells.
- The thin-walled complimentary cells sometimes alternate with bands of dense and compact cells, known as closing cells and the layer formed by these cells is called as a closing layer.

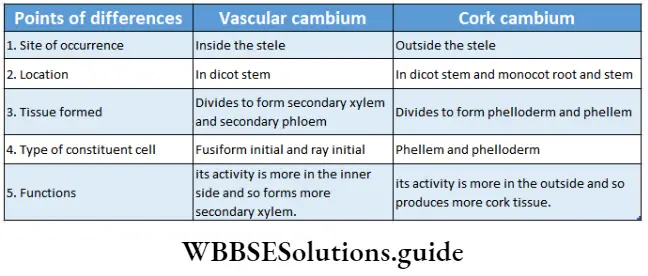
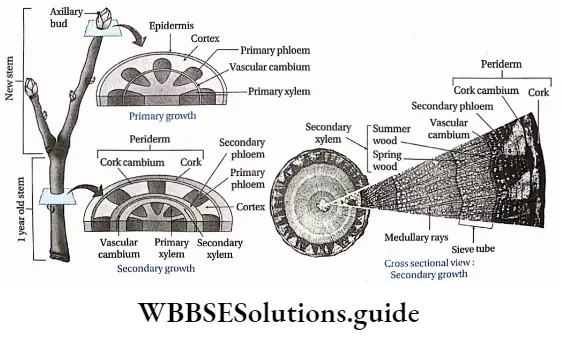
Secondary Growth in Dicotyledonous Root
The secondary tissues formed in the dicotyledonous roots are fundamentally similar to those of the stem, but the process of secondary growth is initiated in a different way. In dicotyledonous roots, secondary growth occurs in intrastellar and extrastellar regions.
Secondary growth in the interstellar region
Formation of cambium ring:
- The dicotyledonous roots have a limited number of radially arranged vascular bundles without any cambium.
- A few parenchyma cells, present below each phloem group, divide and become the meristematic tissue. Thus form cambium strips.
- The number of strips is equal to the number of phloem groups present.
- These cambial cells divide continuously to produce secondary tissues.
- The earlier-formed cambium gradually extends both ways and reaches the innermost cells of the pericycle.
- As a result, a continuous, wavy cambium ring (cylinder) is formed.
Development of secondary xylem and secondary phloem:
- The secondary vascular tissues are basically similar to those of the stem.
- The cambium produces more secondary xylem than secondary phloem. These secondary vascular tissues form a continuous cylinder in which the primary xylem gets completely embedded.
- At this stage, the root structure is revealed only by the radially arranged exarch primary xylem located at the central region.
- The secondary xylem is formed of the tracheid, trachea, xylem parenchyma and xylem fibres.
- Secondary xylem can be easily differentiated from primary xylem as they consist of tracheids with large cavities.
- The secondary phloem is formed of a sieve tube, companion cells, phloem parenchyma and phloem fibres.
- Generally, the primary phloem degenerate. This causes deformation in radially arranged vascular bundles.
- The cambial cells originating from the pericycle opposite to protoxylem groups function as ray initials and produce broad bands of vascular rays.
- These rays developing between the xylem and phloem through the cambium are characteristic of the roots. These rays are called the main medullary rays.
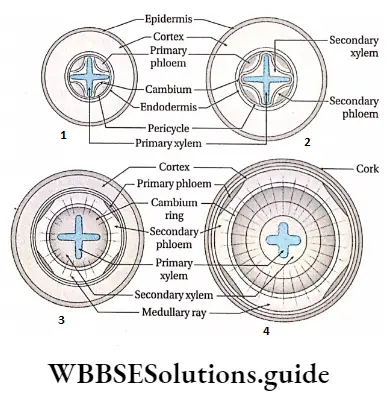
Secondary growth in the extrasolar region:
The secondary growth in the extracellular region occurs in the following phases—
Formation of periderm:
- The cells of the pericycle form phellogen on the outer side of the epiblema. This phellogen or cork cambium develops phellem or cork second cells on the outer side and phelloderm on the inner side.
- Periderm is formed of these three tissues.
- The pressure, exerted by the formation of secondary tissues in the stellar region, ruptures the cortex along with the endodermis.
- Cork is covered with suberin and hence it is impermeable to water.
- It also protects the inner tissues from pathogens.
- It also stores the waste products and helps to release them from the plant body.
Formation of lenticel:
- Some pores are developed between the phellem. These pores are known as lenticels.
- They usually occur in pairs one on each side of a lateral root.
Microscopic Anatomy Of Root, Stem And Leaf
The thin transverse sections of roots, leaves and stems are usually observed under different powers of microscope to study their anatomical features. The different sections are identified through their anatomical characteristics.
The internal structure of the root
- The common identifying features of the root are—
- The epiblema of the root is thin-walled and does not possess a cuticle.
- Epiblema contains unicellular root hairs.
- The endodermal cells have Casparian strips.
- Vascular bundles are arranged radially. A single layer of pericycle is present, from where the branch roots are produced endogenously.
The internal structure of a dicotyledonous root: A transverse section of the root of the leguminous plant—Cicer arietinum (Gram) shows the following arrangement of tissues.
Epiblema or piliferous layer: Epiblema or piliferous layer is an uniseriate outer boundary layer consisting of thin-walled rectangular cells which are longer than their breadth. There are no intercellular spaces between the cells. This layer is devoid of cuticle and stomata. Some cells of epiblema protrude to form long unicellular root hairs.
Cortex: Next to the epiblema, there is a massive but almost homogeneous, parenchymatous zone spread up to the endodermis with conspicuous intercellular spaces. It is referred to as the cortex. The cells are living and contain large amounts of leucoplasts.
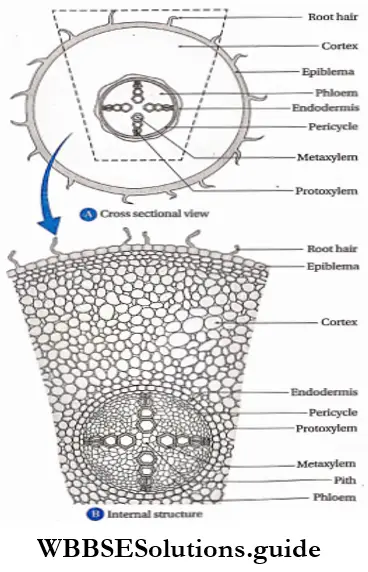
Endodermis: The innermost layer of the cortex is the endodermis, composed of a thin layer of compactly arranged, barrel-shaped cells forming a distinct cell layer around the stele. The cells present in this layer possess characteristic thickenings called Casparian strips on their radial and tangential walls.
Some of the cells of the epidermis, present opposite to the protoxylem, are thin-walled and provide free passage for the diffusion of water and minerals between the cortex and the xylem. They are called passage cells or transfusion cells.
Stele: The central core of tissue, that is surrounded by the endodermis is known as stele. The stele is composed of the following regions—
1. Pericycle: The layer of thin-walled, parenchyma cells without intercellular space situated internal to the endodermis is referred to as pericycle.
2. Vascular bundle: The vascular bundles are radial. In these vascular bundles, the xylem and phloem are arranged on alternate radii. In between the xylem and phloem, small parenchyma cells form a special type of tissue known as conjunctive tissue.
The bundle is tetrarch, as four patches of xylem are arranged alternately with four patches of phloem. Protoxylem vessels are arranged towards the periphery and metaxylem towards the centre. This arrangement is called exarch. A few sclerenchyma cells surrounded each phloem patch.
3. Pith: Usually, pith is not present in dicotyledonous roots. During the early stages of development, small piths made up of parenchyma cells remain situated at the centre, which is later replaced by the development of the metaxylem.
Internal structure of Monocotyledonous Root:
A transverse section of the foot of Colocasia sp. shows the following anatomical features
Epiblema or piliferous layer: The uniseriate epiblema is single-layered, composed of compactly arranged flattened cells without intercellular spaces. Some cells of the layer protrude out to form the unicellular root hairs.
Cortex: The region between the epiblema and the endodermis is known as the cortex. The cortex is mainly formed of parenchyma cells with intercellular spaces. In older roots, cells of the outer layer of the cortex have a suberised wall and form exodermis.
Endodermis: The innermost layer of the cortex constitutes the endodermis consisting of barrel-shaped closely arranged cells with prominent casparian strips. Passage cells or transfusion cells are present opposite to the protoxylem.
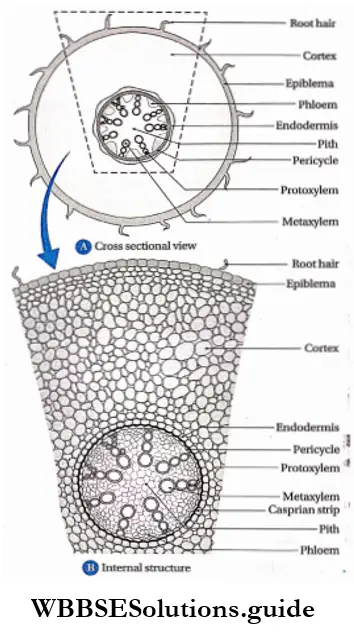
Stele: The central cylindrical core of tissues surrounded by the endodermis forms the stele.
It is composed of radially arranged vascular bundles and interstellar ground tissues.
It consists of the following parts—
- Pericycle: To the inner side of the endodermis, a single-layered parenchymatous cell layer is present, known as the pericycle.
- Vascular bundle: The vascular bundle is radial in nature. A good number of exarch xylem remain alternately arranged with phloem strands. A thin layer of parenchymatous conjunctive tissue separates the xylem and phloem patches. It is polyarchy, i.e., more than six patches of xylem and phloem are present.
- Pith: The central portion of the stele is occupied by a large pith. This region is formed of parenchymatous cells with intercellular spaces.
The internal structure of the stem
- The common identifying features of the stem are
- The epidermis is thick and cuticular.
- Multicellular root hair is present.
- Endodermis bears Casparian strip.
- Pericycle is composed of parenchyma or sclerenchyma
- The vascular bundle is collateral or collateral.
- Xylem is endarch.
- Pith and medullary ray are present.
Internal structure of dicotyledonous stem:
When a transverse section of the stem of a sunflower (Helianthus annuus) is observed under a microscope, the following tissues are seen (serially from the periphery)—
Epidermis: It consists of a single layer of barrel-shaped parenchymatous cells, without any intercellular spaces between them. Multicellular shoot hairs originate from this layer. A distinct noncellular covering made of cutin is present as the outermost layer called the cuticle.
Cortex: The region between the epidermis and endodermis is the cortex.
This region is divided into three parts—
Hypodermis: This region is made up of 4-5 layers of living collenchyma tissue.
General cortex:
- It is the middle region between hypodermis and endodermis which consists of several rows of parenchyma cells.
- The resin duct is scattered irregularly in this layer.
Endodermis:
- This layer is wavy and made up of a single layer of barrel-shaped parenchyma cells.
- Due to the presence of starch granules, this layer is also called starch sheath.
Stele: Stele is formed of the following layers—
Pericycle:
- Pericycle is made up of both parenchymatous and sclerenchymatous cells.
- The parenchyma cells form a continuous outer layer. Inside which the sclerenchymatous layer lies.
- Above each vascular bundle, several layers of sclerenchyma cells exist like a cap or sheath. They are known as bundle sheaths.
- The sclerenchymatous layer is interrupted by medullary rays.
Vascular bundle:
- This vascular bundle is conjoint, collateral and open,
- The upper part of each vascular bundle bears a phloem while the lower part bears a xylem. As cambium is present between the xylem and phloem, the vascular bundle is open.
- Medullary rays: Thin-walled radially elongated parenchyma that emerges from the pith in the form of rays between two vascular bundles is called medullary rays.
- Pith: This part of the stem is either oval or spherical and is composed of parenchyma cells.
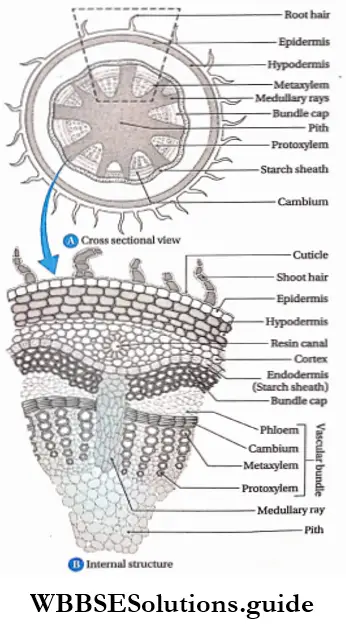
Internal structure of monocotyledonous stem: When a transverse section of the stem of maize (Zea mays) is observed under a microscope,
The following tissues are seen (serially from the periphery)—
Epidermis:
- It is formed of a single layer of barrel-shaped parenchyma cells.
- The epidermis possesses a cuticle.
- Parenchyma cells are filled with chlorophyll. The outer surface does not bear any hair.
Hypodermis: This layer is present beneath the epidermis and is composed of 2-3 layers of scarified parenchymatous cells.
Ground tissue:
- This tissue extends from the hypodermis to the centre. It is composed of thin-walled parenchyma cells. Cells have intercellular spaces.
- Vascular bundles remain scattered in this region.
- The Stem of Lea Mays does not have endodermis, pericycle or pith.
Vascular bundle:
- The vascular bundle is conjoint, collateral and closed.
- At the periphery of the stem, the vascular bundle becomes smaller and more in number. At the centre, they are larger and fewer in number.
- Each vascular bundle is surrounded by sclerenchyma. This is known as a bundle sheath.
- The vascular bundle has only the xylem and phloem. Xylem is arranged as T within which phloem is present.
- The arms of Y represent the metaxylem and the leg is the protoxylem.
- The lowermost cavity of the protoxylem is called the protoxylem cavity or lysigenous cavity. As the vascular bundles remain scattered in ground tissue, there is no pith at the centre.
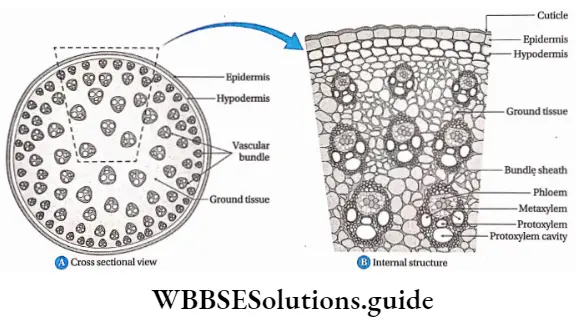
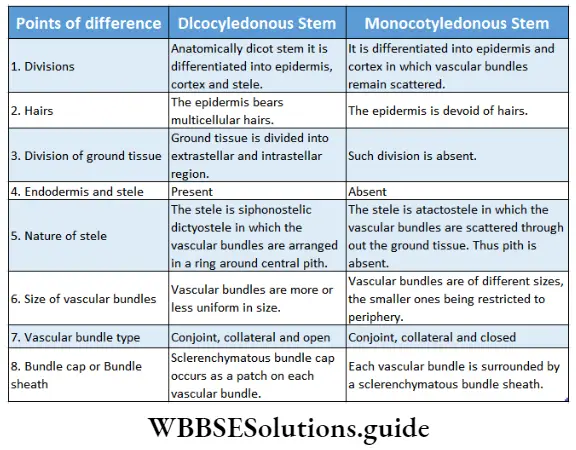
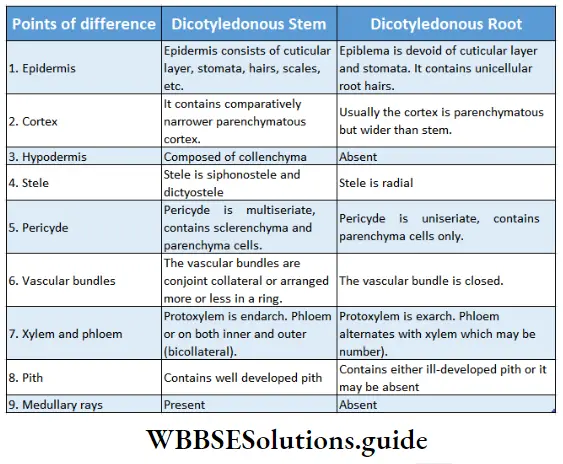
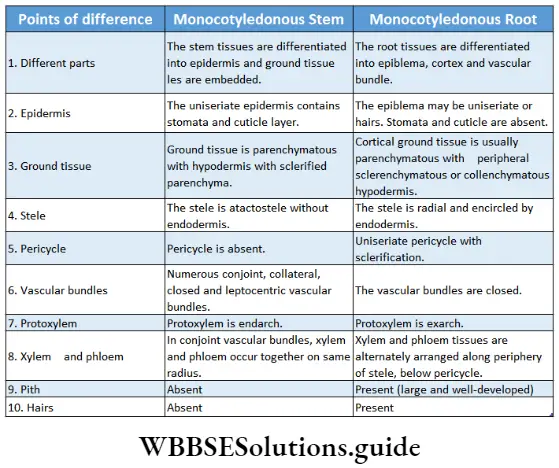
The internal structure of Leaf
The common identifying features of the leaf are:
- Anatomically the leaves are composed of different tissue systems.
- The epidermal tissue system contains epidermal layers on both the upper or adaxial and lower or abaxial sides with stomata and outgrowths.
- In the leaf, the ground tissue system, the mesophyll tissue, is present. Usually, it is differentiated into columnar palisade parenchyma on the adaxial side and isodiametric or irregularly shaped spongy parenchyma on the abaxial side.
- Mesophyll tissue is provided with conspicuous air spaces, that help in the gaseous exchange with the atmosphere.
- Vascular bundles are closed and collateral in nature.
- Xylem is research.
- Stomata is always present on the abaxial surface of the leaves.
Stomatal Density and Stomatal index
The distribution of stomata is an important feature of plants. It varies between the upper and lower epidermis and between dicotyledonous and monocotyledonous plants. It varies with changes in environmental factors like sunlight, CO2, humidity, etc.
This can be studied by removing the peels of the upper and lower surfaces of the leaf, with forceps and observing under a microscope.
The number of stomata and epidermal cells per mm2 of leaf surface area is taken into account. The stomatal density and stomatal index can be calculated by the following formulae:
Stomatal density (SD)= Number of stomata per mm2 Stomatal index (SI)= (SD x 100)/(SD + Number of epidermal cells per mm2).
On the basis of anatomical features, leaves are of three types—
- Dorsiventral or bifacial leaves;
- Isobilateral or equifacial leaves and
- Unifacial leaves.
The internal structure of the dorsiventral or bifacial leaf: Dorsiventral leaves are arranged horizontally to the ground. Because of unequal exposure to sunlight on the two sides, they have distinct upper and lower surfaces. Most of the dicot plants have dorsiventral leaves.
A vertical section through the leaf lamina of the mango shows the following arrangement of tissues.
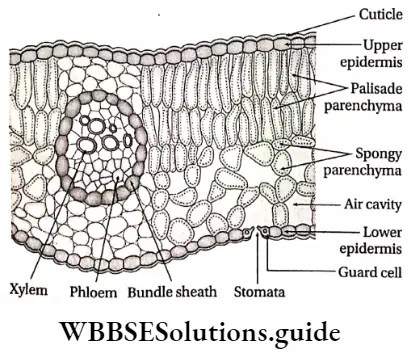
Epidermis: The epidermal layer is formed of living barrel-shaped parenchymatous cells. This layer is divided into two parts—the upper epidermis and the lower epidermis. The outer walls of epidermal cells possess a thin cuticle.
Upper epidermis: The upper epidermis has a thicker cuticle.
Lower epidermis: Stomata occur on the lower epidermis, thus the leaf is hypostomatic.
Mesophyll: The region between the upper and lower epidermis is ground tissue’ known as mesophyll tissue. The mesophyll tissue is composed of two types of parenchyma cells, palisade and spongy parenchyma.
Palisade parenchyma: The columnar palisade parenchyma cells, with fewer intercellular spaces, are present beneath the upper epidermis. These oblong-shaped cells remain at right angles to the leaf surface. Palisade cells contain a large number of chloroplasts along the peripheral walls. There are two layers of palisade cells.
Spongy parenchyma: The spongy parenchyma cells occur adjacent to the lower epidermis. They are very loosely arranged with large intercellular spaces. These cells contain a comparatively lesser number of chloroplasts. Hence, the lower surface of the leaf is pale green in colour.
Vascular bundles:
- The position of the vascular bundle depends on the type of leaf venation. In these leaves, vascular bundles are present at the connecting point of the palisade and spongy parenchyma.
- The vascular bundles are mesarch, collateral and closed.
- The size of the vascular bundle varies depending on its position in the leaf.
- A larger vascular bundle is composed of an xylem situated towards the upper epidermis and a phloem towards the lower side.
- Individual vascular bundle remains encircled by bundle sheath.
- Parenchyma or collenchyma cells connect the bundle sheath with two epidermal layers, called bundle sheath extension.
The internal structure of the isobilateral leaf:
Isobilateral leaves are oriented angularly to the ground, thus both upper and lower surfaces are equally exposed to sunlight. These leaves possess uniform structural organisation on both surfaces. Most of the monocot plants have isobilateral leaves.
A transverse section through maize (Zecr mays) leaf blade shows the following anatomical features.
Epidermis: This layer is formed of parenchyma cells with intercellular spaces. The epidermis is divided into two layers—the upper epidermis and the lower epidermis.
Both the epidermal layers (upper and lower) are uniseriate and composed of more or less oval cells. A cuticle layer is present on the leaf surface. Stomata are present on both the epidermal layers, thus the leaf is amphistomatic.
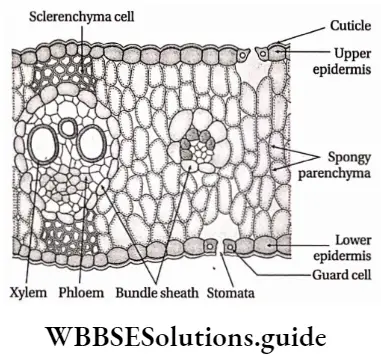
Upper epidermis: This layer is formed of oval-shaped, closely arranged parenchymatous cells. The outer wall is uniformly cuticularised. The upper epidermis contains large and empty bulliform cells. A large number of stomata are present in this layer.
Lower epidermis: It is cuticular like the upper epidermis but does not contain any bulliform cells. This layer also contains the same number of stomata as present in the upper epidermis.
Mesophyll: The mesophyll tissue is not differentiated into palisade and spongy cells. It is composed of compactly arranged, isodiametric cells. In these leaves, all the mesophyll cells are spongy types.
Vascular bundles: The vascular bundles are arranged in parallel lines, and they are collateral and closed in nature. Most of the vascular bundles are small. Larger bundles occur at regular intervals.
Each vascular bundle has a xylem on the upper side and a phloem on the lower side, surrounded by a sclerenchymatous bundle sheath.
Anatomy of flowering plants summary with important points
The bundle sheath cells contain plastids which are without grana (agranal) or with a few grana filled with starch grains. Sclerenchyma cells occur in patches on both edges of the bundles external to the bundle sheath. These cells provide mechanical strength to the leaves.
Unifacial leaves
Unifacial leaves develop from one side of the leaf primordia and have only an encircling adaxial or abaxial epidermis. Some plants have cylindrical leaves with no distinction into an upper or lower surface (for example onion) or flattened (for example, mint).
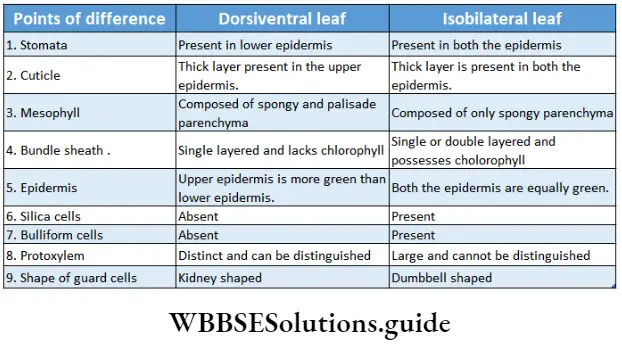
Anatomy Of Flowering Plants Notes
- Caspian strips: In root endodermal cells possess bands of thickening along the radial or tangential walls. These are called caesarian strips or Caspian bands. They are made of lignin and suberin. They prevent the plasmolysis of endodermal cells and prevent the movement of substances between the cortex and pericycle.
- Cutin: It is a waxy polymer associated with the cell wall, found in plants. This forms the cuticle of the epidermis of different plant parts.
- Drupe: A type of fleshy single-seeded fruit. Mango and coconut are examples of drupe.
- Ergastic substances: These are some products of cell metabolism found either in vacuoles or cytoplasm. These include gums, tannins, oil droplets, resins, etc.
- G0 Phase: This is the period of the cell cycle when cells neither divide nor prepare to divide, but rather survive.
- Hemicellulose: It is a complex polysaccharide, a chief building material of the cell wall.
- Latex: Latex is the secretion of latex cells. It is usually yellow or white, milky or watery fluid.
- Leafgap: The leaf gap is a break in the vascular tissue of a stem just above the point of extension of a strand of conducting vascular bundle from the stem to the leaf base.
- Lignin: It is an organic polymer, associated with a cell wall. It is a main component of wood and bark. it is a water-resistant substance that gives the cork its impervious nature.
- Lithocyst: A large epidermal cell, that contains a large calcium carbonate crystal (cystolith) on an ingrowth of the cell wall.
- Ontogeny: All the developmental events that occur during an organism’s life. This begins with changes in the egg at the time of fertilization and includes all the developments that take place up to the time of birth and afterwards.
- Pectin: It is a cell wall-associated polymer, a major component of the primary cell wall of the terrestrial plant.lt helps in the formation of wood.
- Phytogeny: The history of evolution of a species especially in reference to the line of descent and relationship among different taxa of organisms.
- P-protein: A protein found in phloem tissue. In injured or disrupted sieve elements, they aggregate at the sieve plate to plug the leakage of phloem exudate.
- Primordial utricle: In a fully developed plant cell, cytoplasm moves towards the membrane forming a thin layer surrounding the large central vacuole. This cytoplasmic lining is called the primordial utricle.
- Suberin: It is an inert impermeable waxy polymer found in the cell walls of woody plants. It is water resistant and a major constituent of cork.
- Tension wood: In woody angiosperms, this high cellulose-containing type of wood is formed as a response to gravity, where cambium is not vertically positioned. It is. typically found in branches and leaning stems.
- Testa: In a dicot seed, the outer seed coat is called testa.
Points To Remember
- In plant physiology, the term ’tissue’ was first used by N. Grew (1682).
- On the basis of divisional property, plant tissue has been divided into two types—
- Meristematic tissue and
- Permanent tissue.
- The tissue whose cells are bound by a thin membrane and divide by mitosis to give rise to new cells is called meristematic tissue.
- The term ‘meristem’ was first coined by Nageli (1858).
- The meristematic tissue that develops directly from embryonic cells is called primary meristematic tissue and the meristematic tissue that develops from permanent tissue is called secondary meristematic tissue. Cork cambium, interfascicular cambium and cambium present in plant roots are examples of secondary meristematic tissue.
- On the basis of the plane of cell division, meristematic tissue is divided into three types—mass meristem, plate meristem and rib meristem.
- On the basis of location, meristematic tissue is divided into three types—
- Apical meristem,
- Intercalary meristem,
- Lateral meristem.
- On the basis of function, meristematic tissue is of three types—
- Protoderm (the outermost layer of apical meristem from which epidermis and epiblema develop).
- (Procambium (from which primary vascular tissue develops).
- Ground meristem (present beside epidermis and vascular tissue system, from which other tissue systems develop).
- The inactive centre at the root apex just below the root cap region contains lesser DNA, RNA and proteins and is called the quiescent centre.
- Based on the types of cells involved in composition, permanent tissue is divided into three types—
- Simple permanent tissue,
- Complex permanent tissue and
- Special permanent tissue.
- Simple permanent tissue is of three types—
- Parenchyma,
- Chollenchyma And
- Sclerenchyma
- Parenchyma cell contains many excretory substances like tannin, resin, calcium oxalate, crystal, benzoin resin, etc.
- Cell walls of collenchyma cells possess cellulose and pectin.
- Collenchyma cells are divided into three types, on the basis of the thickness of the cell wall—
- Angular,
- Lacunar and
- Lamellar. Stratified collenchyma is also called plate collenchyma.
- On the basis of shape and size, sclerenchyma is divided into two types—sclerenchyma fibres and chloride.
- Sclerenchyma fibres are elongated and have pointed ends.
- Sclerenchyma fibres are divided into two types, intraxylary (the cells of sclerenchyma that are present within the xylem) and extraxylary (the cells of sclerenchyma that are present outside the xylem i.e., within cortex, pericycle and phloem fibres).
- The type of sclerenchyma tissue in which constituent cells are spherical, columnar and irregular in shape and whose cell wall is thickened by lignin, suberin and cutin, is called scleride. Since the cells have the same radius, tough cell wall and gritty texture. These cells are called stone cells.
- Xylem fibres are called wood fibres and phloem fibres are called bast fibres.
- Living phloem parenchyma store food and dead phloem fibres provide mechanical support to the plant.
- Xylem in roots are exarch in nature.
- The vascular bundle in which cambium is not present between the xylem and phloem is called a closed vascular bundle.
- The vascular bundle in which cambium is present between the xylem and phloem is called an open vascular bundle.
- The stele which lacks pith is called protostele. example Lycopodium.
- The method by which the age of a tree is determined by counting its annual rings is called dendrochronology.
- The multicellular, hair-like epidermal appendage in plants that helps in secretion is called trichomes.
- The wood produced by cambial tissue under adverse conditions in winter is called autumn wood or latewood.
- The central, hard part of the dicot stem filled with tannin, resin, etc., is called heartwood or duramen.
- Phellem, phellogen and phelloderm fuse to form periderm.
- The Endodermis of the root possesses passage cells just adjacent to the xylem.