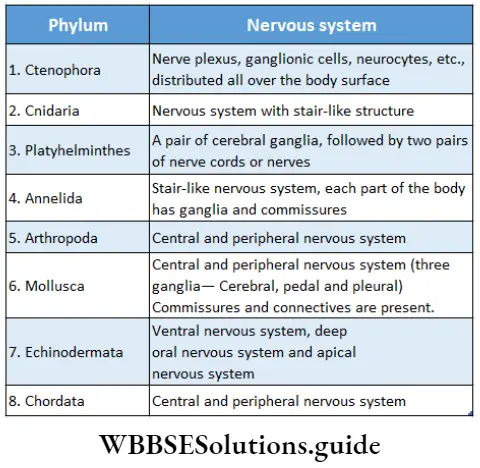
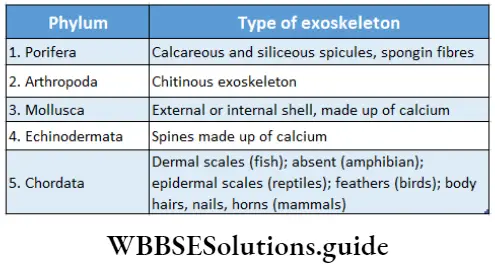
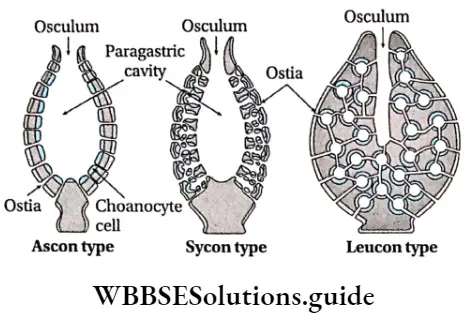
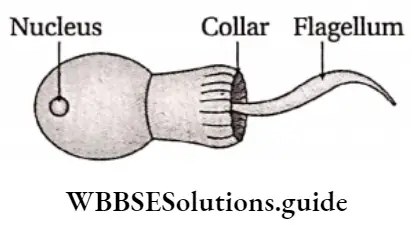
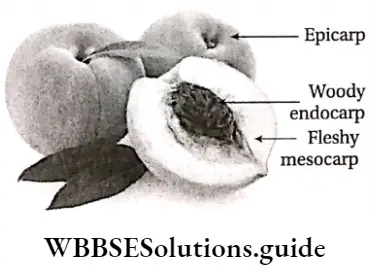
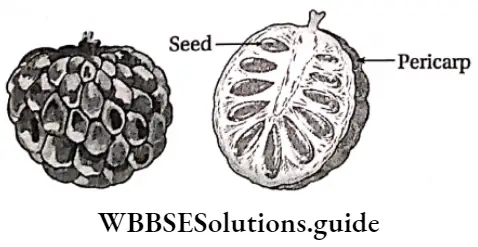
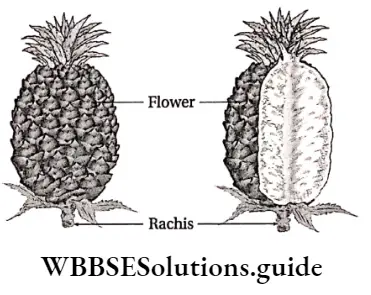
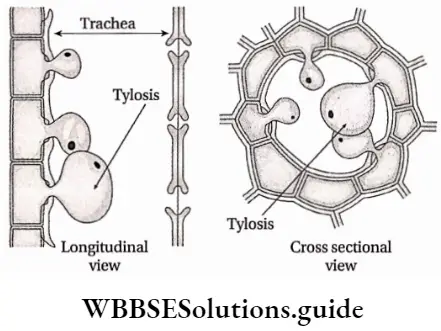
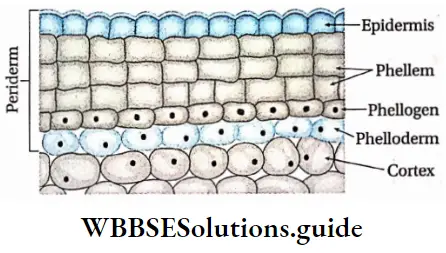
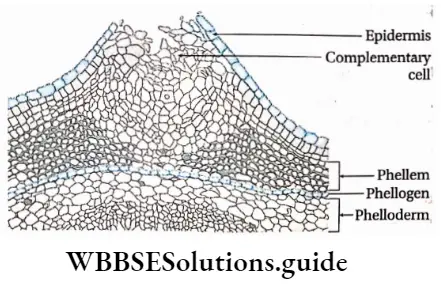
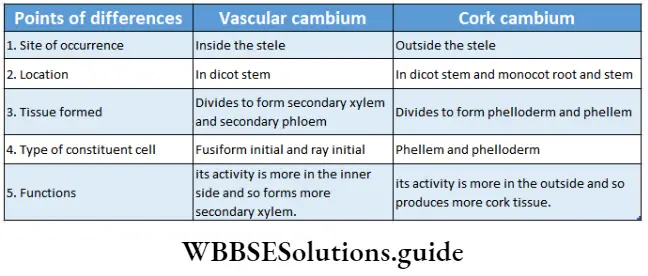
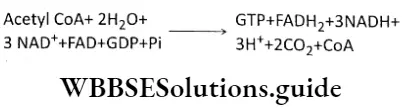
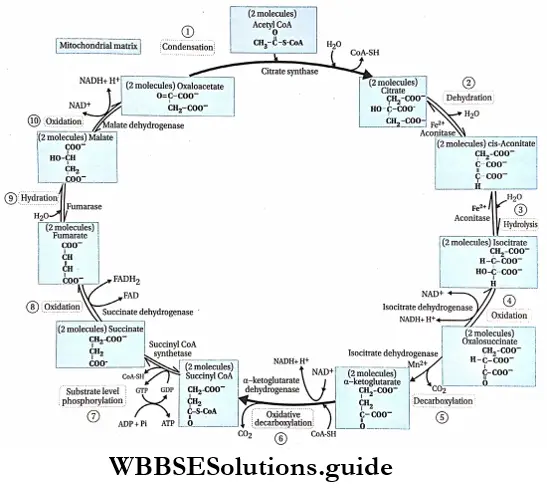
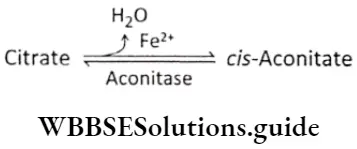
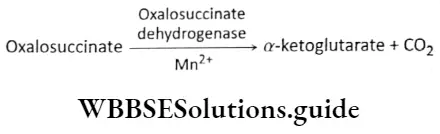
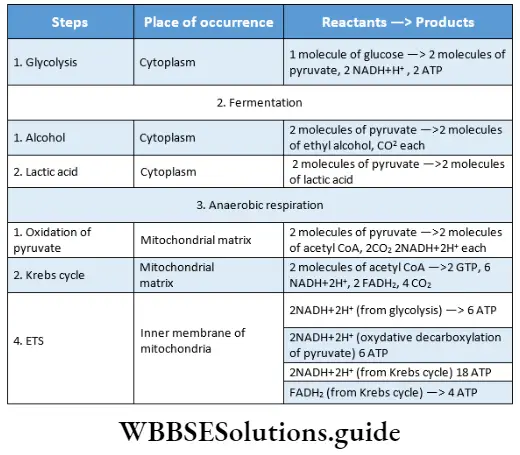
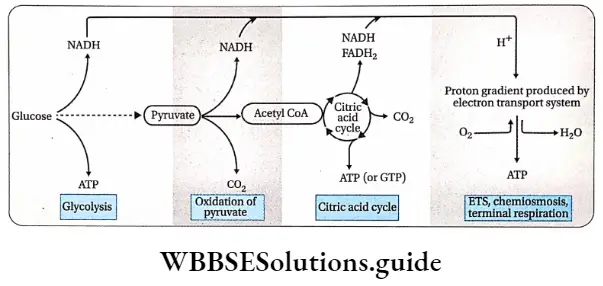
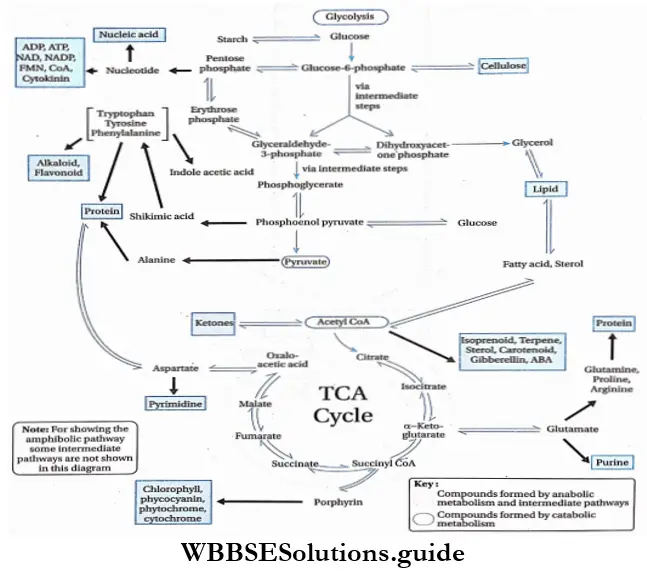
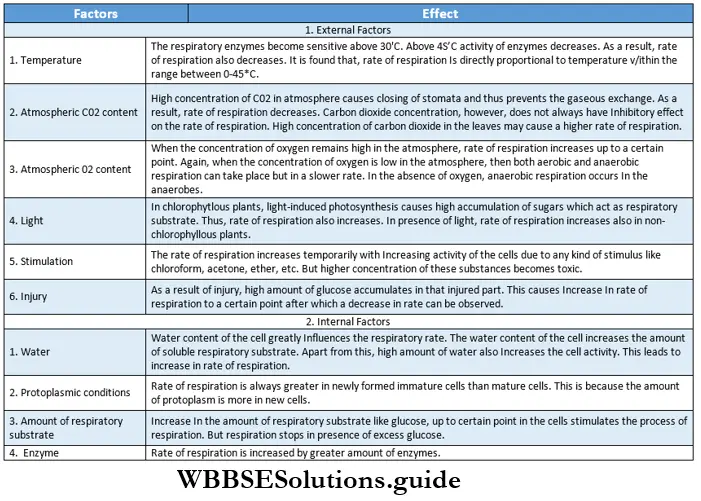
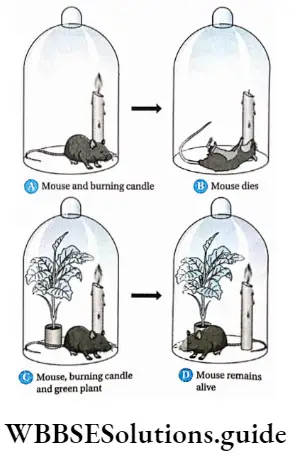
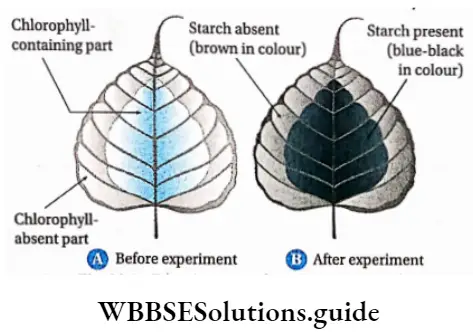
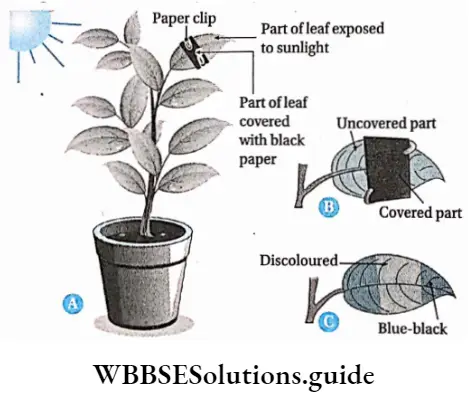
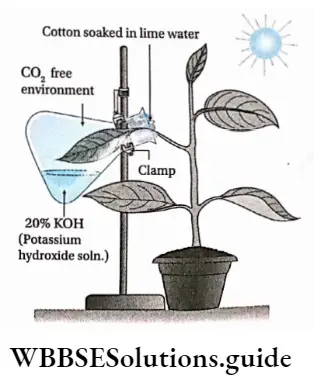
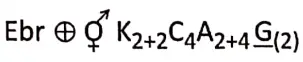Plant Kingdom Introduction To Play Kingdom
The kingdom Plantae or plant kingdom includes all eukaryotic chlorophyll-containing autotrophic organisms which are commonly called plants. Some of the general features of the plants are as follows
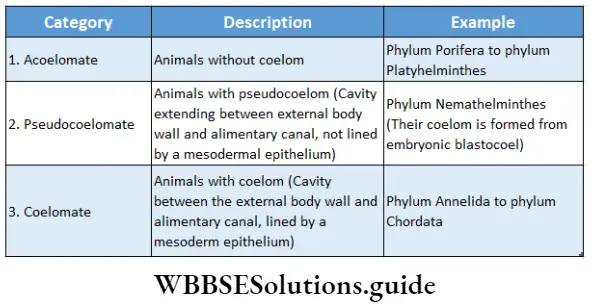
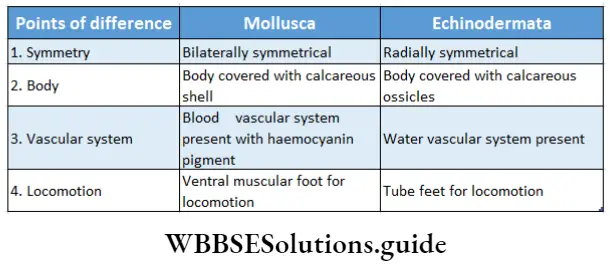
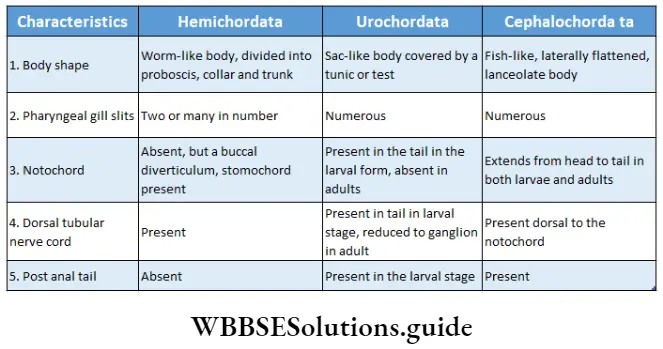
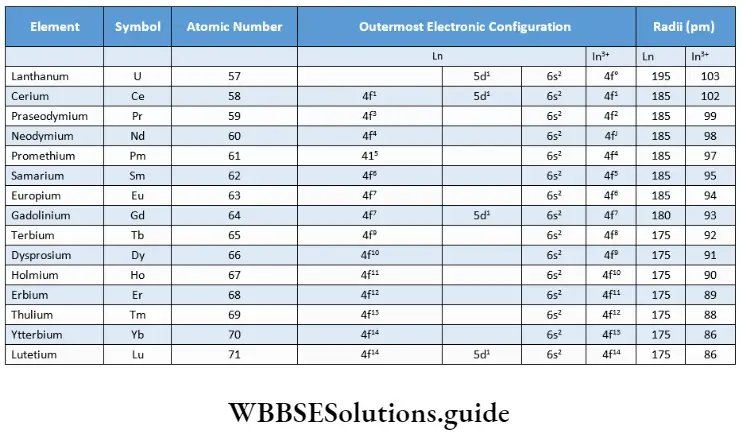
Habitat: This kingdom comprises the following groups—Algae, Bryophyta, Pteridophyta, Gymnosperms and Angiosperms. The algae thrive in aquatic habitats. Members of the Bryophyta and Pteridophyta include both aquatic and terrestrial members.
The gymnosperms and angiosperms are mostly terrestrial. However, they also include several aquatic species as well, for example, Hydrilla sp., Vallisneria sp., etc.
The aquatic species are found in freshwater, brackish water or seawater. The terrestrial members may live in different places such as mountains, deserts, marshlands, river basins, etc.
Read and Learn More: WBCHSE Notes for Class 11 Biology
Structural organisation: All the members of this kingdom are eukaryotic in nature. Many members of primitive groups, i.e., algae and bryophytes, have a thallus-like body structure. Such a body is not differentiated into roots, stems and leaves.
The body of the higher plants, i.e., pteridophytes, gymnosperms and angiosperms, are generally differentiated into true root, stem and leaves.
“plant kingdom classification and characteristics notes for class 11”
Nutrition: Most plants are autotrophic in nature, they can synthesise their own food. Some may be partially heterotrophic such as insectivorous plants or parasitic plants. Bladderwort and Venus flytrap are examples of insectivorous plants. Cuscuta sp. is an example of a parasitic plant.
Cell covering and vacuoles: The plant cells have a eukaryotic structure with a prominent cell wall. The cell wall is mainly made up of cellulose. Each mature cell may have a large central vacuole.
Plastids: All plants have plastids in their cells. The green parts of the plant have chlorophyll-containing chloroplastids. The coloured parts have chromoplastids, that contain pigments other than chlorophyll. The parts of the plant that store food, contain leucoplastids, which are without any pigment.
| Class 11 Biology | Class 11 Chemistry |
| Class 11 Chemistry | Class 11 Physics |
| Class 11 Biology MCQs | Class 11 Physics MCQs |
| Class 11 Biology | Class 11 Physics Notes |
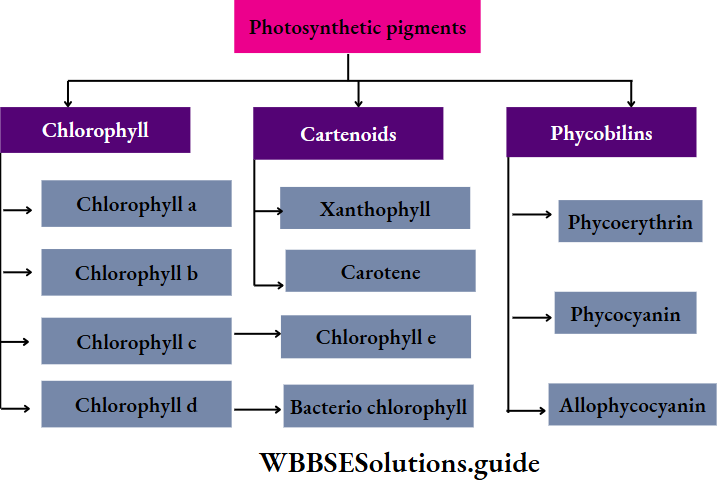
Pigments: Chlorophyll a is present in the chloroplasts of almost all plants. Chlorophyll b, c, and d are present in addition to Chlorophyll a, in different plant groups. Other than chlorophyll, carotenoids (orange), xanthophyll (yellow), phycocyanin (blue-green), phycoerythrin (red), etc., are also present.
Reserve food: Starch is the major reserve food in plants. Other than this, fat or oil droplets, some proteins and crystals of different salts are also found in plant cells as reserve food.
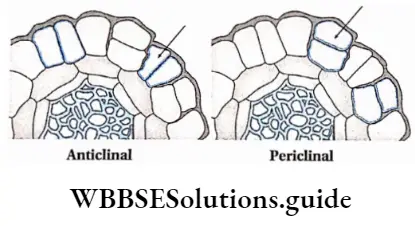
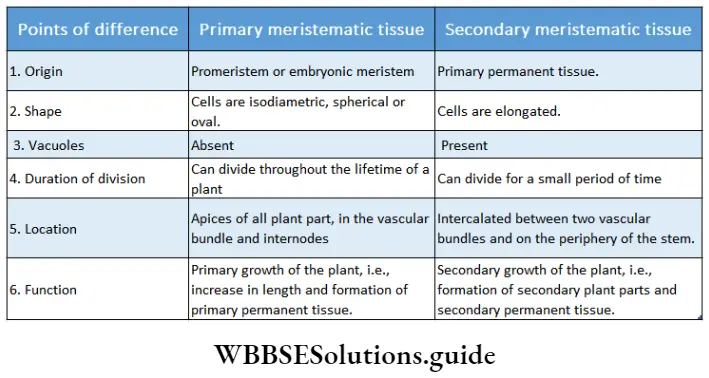
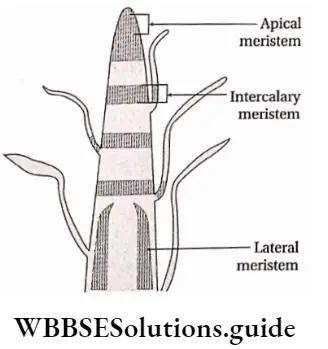
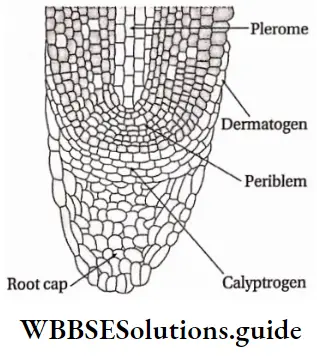
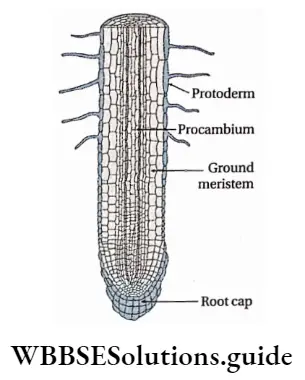
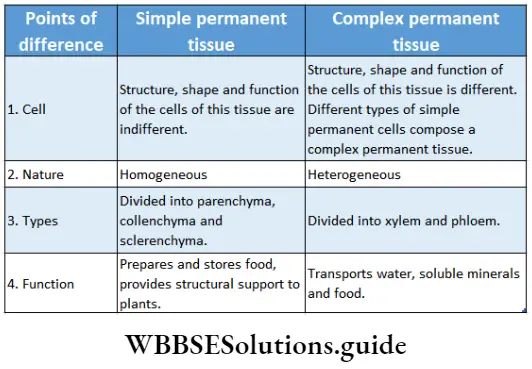
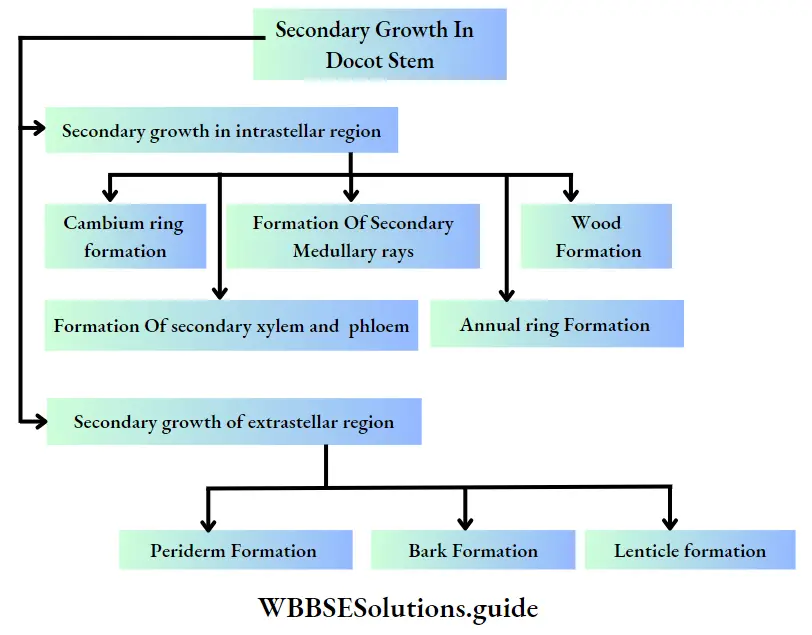
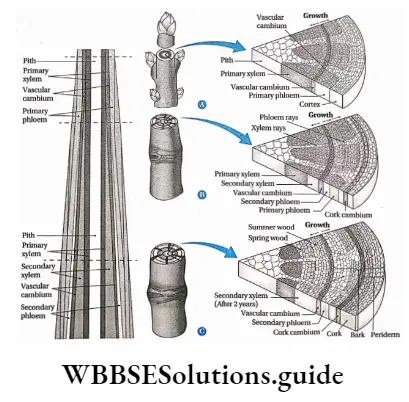
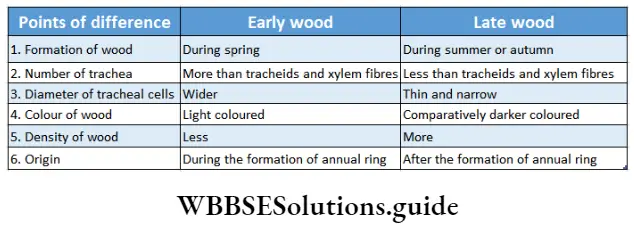
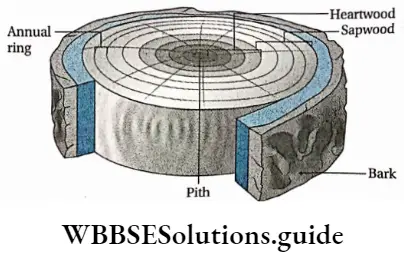
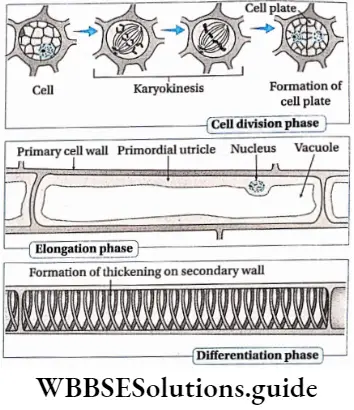
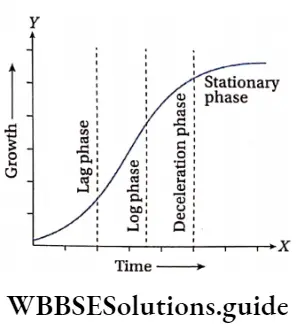
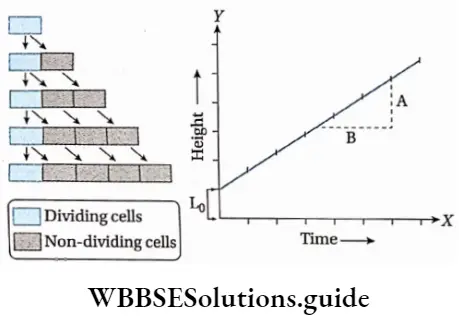
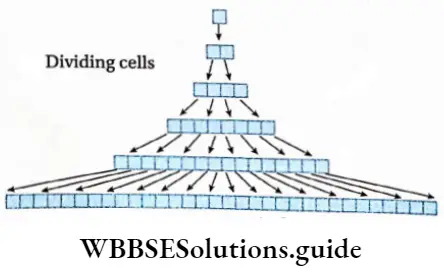
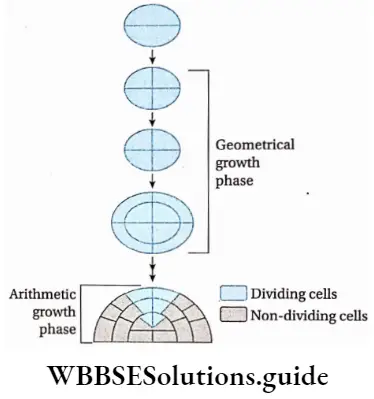
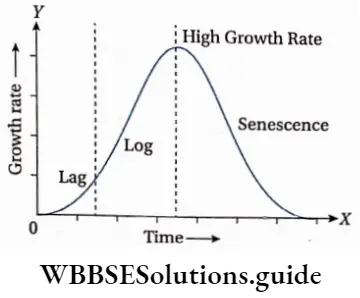
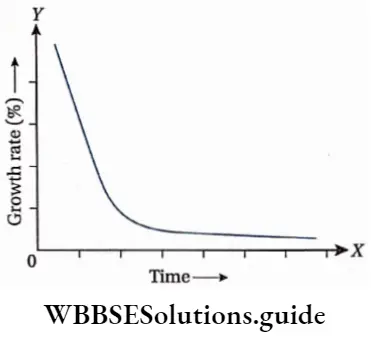
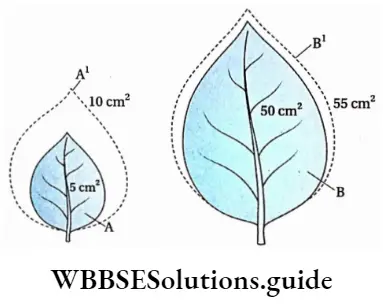
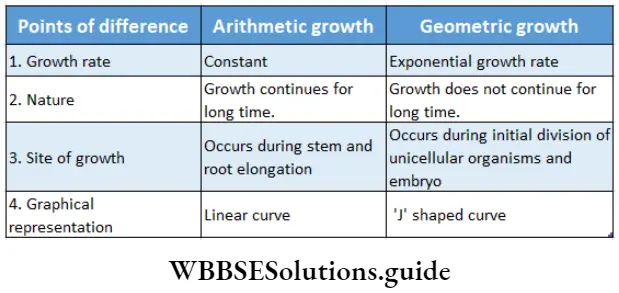
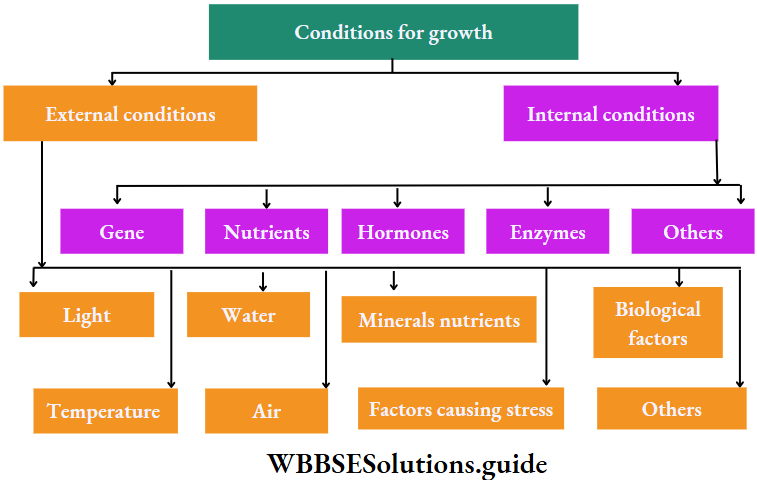
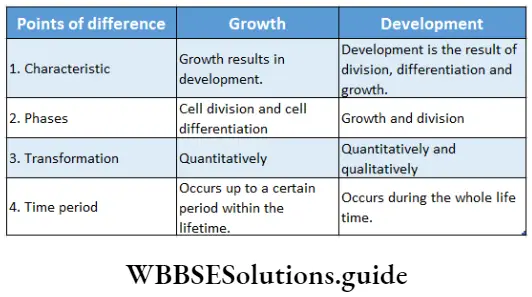
Growth: Except in some unicellular plants, all other plants show unregulated growth. Meristematic tissues are the tissues that form various plant parts and keep the plant growing. Some higher plants also show secondary growth besides primary growth.
“detailed notes on plant kingdom classification and characteristics”
Movement and locomotion: Only some unicellular green algae are able to show locomotion. The rest of the plants are static in one place and spread their branches from there. However, some plants (such as algae, bryophytes, pteridophytes and some gymnosperms) produce motile gametes (reproductive units or cells).
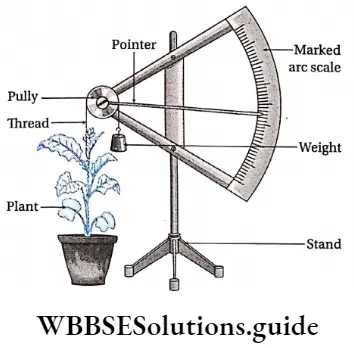
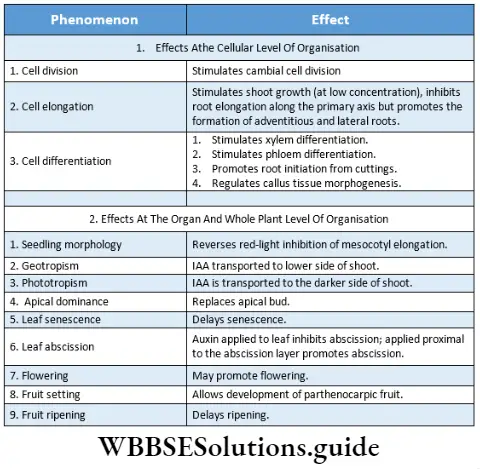
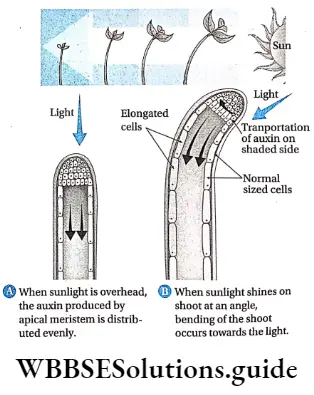
Response to stimuli: Despite the absence of a nervous system, plants respond to external stimuli. They produce several growth-regulating chemicals to respond to environmental stimuli. Movement or growth toward stimuli like gravity, light, water, etc., is known as tropism. Shoots tend to grow or move toward light source (phototropism) and roots grow towards gravity (geotropism).

The male gametes of bryophytes show chemotropism (stimulus for such movement is a specific biochemical substance) towards female gametes.
” kingdom plantae examples”
If the response to stimuli is independent of the direction of stimuli, then such response is called nastic movement. Insectivorous plant Venus flytrap rapidly closes its laminar halves when an insect sits on its leaf.
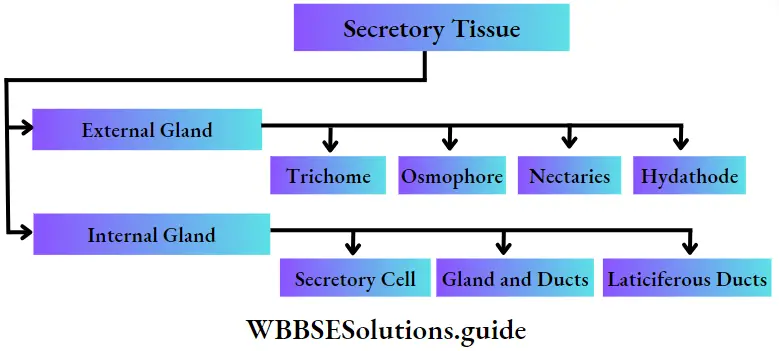
Presence of vascular tissues: Except for algae and bryophytes, all plants have vascular tissues to transport water and nutrients across the cells. In some bryophytes, there are special tissues, called hydroid and leptoid, that are similar to vascular tissues.
“plant kingdom classification with examples and characteristics”
Reproduction: Reproduction in plants is of three types—vegetative, asexual and sexual. Plants reproduce asexually by different types of asexual reproductive units or spores. In angiosperms, vegetative reproduction occurs through underground stems, roots, leaves, etc.
During sexual reproduction, plants produce male and female gametes. These gametes may or may not be morphologically similar Union of gametes leads to the formation of zygotes which develop into new plants.
Alternation of generations and life cycle: Life cycle patterns of the members of the plant kingdom show two distinct phases—the diploid (2n) or sporophytic phase and the haploid (n) or gametophytic phase—that alternate with each other throughout the life cycle.
This phenomenon is called the alternation of generations. The duration of each phase, and their dependence on each other, vary, among different groups of plants.
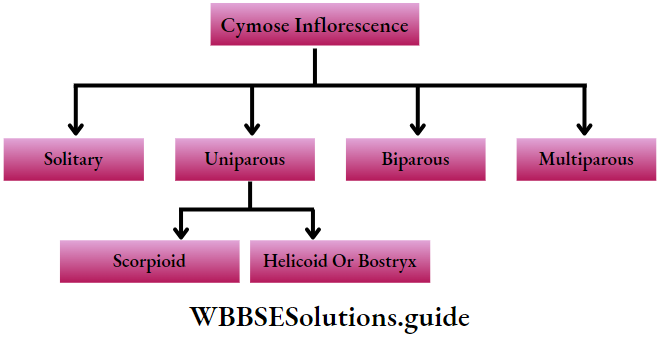
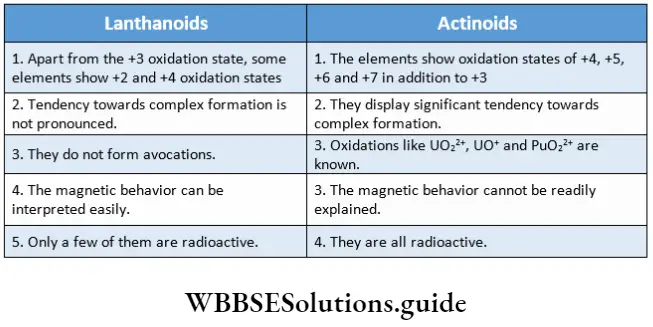
Classification Of Plant Kingdom Into Major Groups
The history of plant classification is fascinating. To understand the concerns that affected the classification system let us consider classification within Angiosperms.
Classification is a logical system of organisation of several categories or taxa, each containing one or more organisms. Plants are also classified into different taxas by different authors. These classifications are categorised into three types—artificial, natural and phylogenetic.
Artificial system: It is the earliest system of classification of plants, which was based on one or a few morphological features such as habit, colour, number ar|d shape of leaves, floral characters, etc.
” biological classification of plants “
Theophrastus (370-285 B.C.), the father of botany,c|assjfjed a|| p|ants on the basis of form and texture. John Ray (1628-1705) broadly divided plants into two groups -Herbae, i.e., herbaceous and Arborae, i.e. woody. The Herbae were divided into two groups Imperfectae (non-flowering plants or cryptogams) and Perfectae (flowering herbaceous plants).
The group Arborae contains the flowering trees. He classified the group perfectly and airborne into monocots and dicots. Carolus Linnaeus (1936), also known as the father of taxonomy classified the then existing plants based on number, union and length of stamens.
Natural System: In this system, the plants are grouped on the basis of both external and internal features, like ultrastructure, anatomy, and embryology. Classification propounded by George Bentham and sePh Dalton Hooker is one of the most popular natural systems of classification.
This was published in the book Genera Plantarum (1862-1883). In Bentham and Hooker’s system of classification, plants are grouped into two sub-kingdoms, Cryptogamia (non-flowing plants) and Phanerogamia (flowering plants).
Phylogenetic system: In this system, plants are classified on the basis of their course of evolution, and genetic and phylogenetic relationships. Some examples of a phylogenetic system of classifications are those propounded by A. W. Eichler (1875), A. Engler and Karl A. E. Prantl (1936), John Hutchinson (1959), etc.
In some modern phylogenetic systems of classification information available from various fields of study, such as palaeobotany, biochemistry, cytology, molecular biology, anatomy, etc., are used along with morphological features of the plants.
“kingdom plantae classification “
Classifications propounded by A. Takhtajan (1980), Arthur Cronquist (1981) and Thorne (1983) are examples of modern phylogenetic systems.
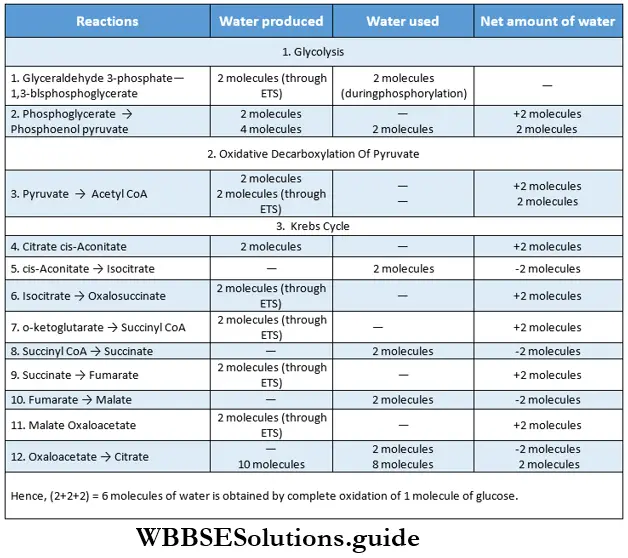
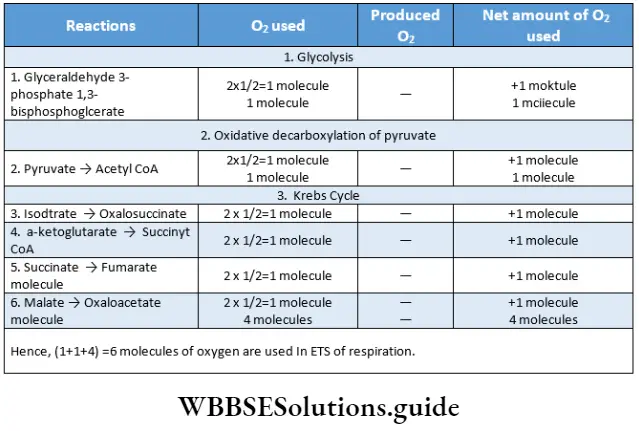
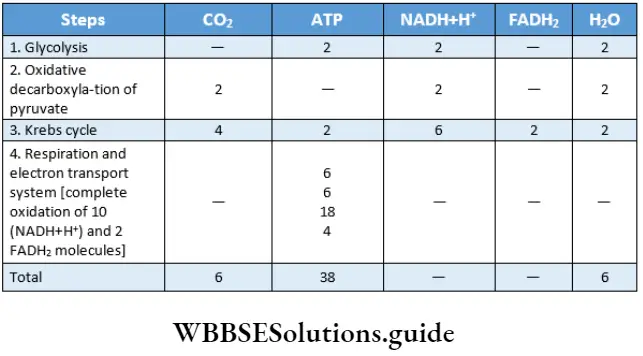
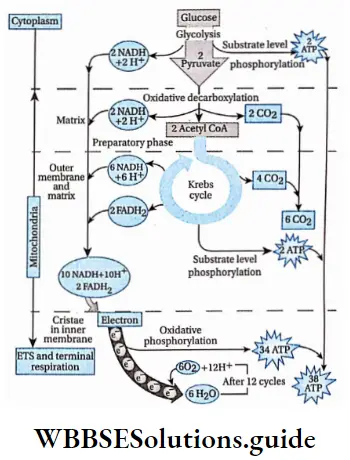
The outline classification of plants which is widely accepted among the biologists is shown below in tabular form.
Each group mentioned in this classification has been discussed separately, later in this chapter.
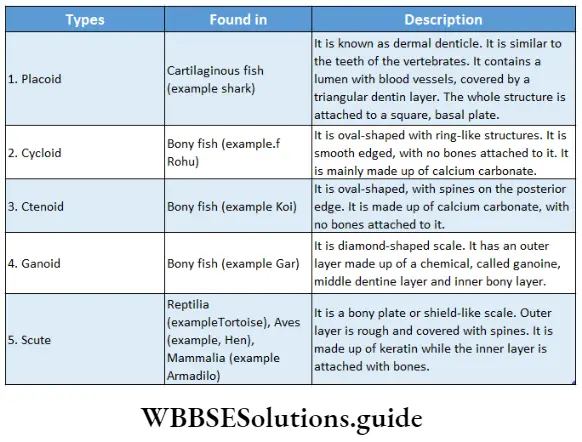
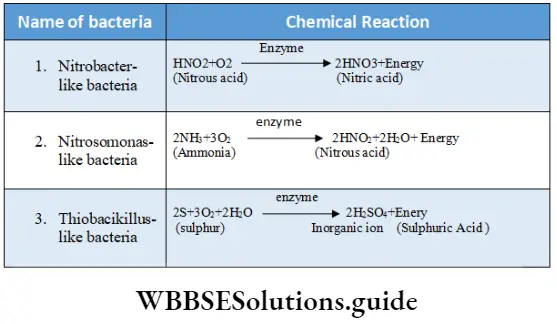
Phaeophyceae (Brown algae)
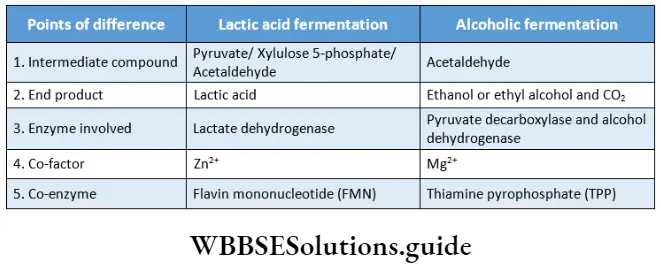
Phaeophyceae Definition: Phaeophyceae (Greek: phaios= brown; phyton = plant) are the multicellular, mainly marine, brown-coloured group of algae.
This group of algae is commonly known as brown algae, as they appear golden brown in colour due to the presence of a xanthophyll pigment—fucoxanthin. The fucoxanthin partially masks the chlorophylls and carotenes. Thus gives the characteristic brown colouration.
It is a large group of algae consisting of 240 genera and over 1500 species. Among these 32 genera and 93 species have been reported from India.
“biological classification of plants “
Phaeophyceae Distribution: They are mostly marine in habitat and a few are freshwater. The freshwater members are Pleurocladia spv Heribaudiella sp., Lithoderma sp. and Sphacelaria sp. Pleurocladia lacustris grow both in freshwater and marine habitats. They are generally found attached to the marine substratum, in the colder regions of the sea.
Phaeophyceae General features:
- They range from simple microscopic (1 mm) heterotrichous filament (Ectocarpus sp.) to the largest alga (60-90 metres) (Macrocystis pyrifera).
- Giant brown algae are called kelps. The plant body is non-motile, multicellular and highly differentiated, both externally and internally. Their shapes may range from ribbon-like, filamentous, leaf-like, bush-like, fern-like or some may be branched like trees. Unicellular, colonial and unbranched filamentous forms are completely absent.
- The plant body is differentiated into holdfast, short or elongated stipe and an expanded leaf-blade or lamina. The lamina performs photosynthesis and bears reproductive structures. Many species can float due to the presence of air bladders.
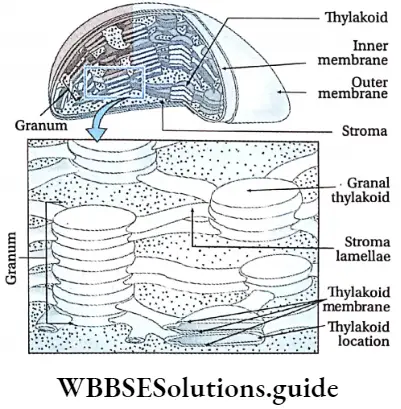
- The photosynthetic plastids present in Phaeophyceae are called phaeoplasts. The phaeoplasts contain three thylakoid-containing lamellae. The photosynthetic pigments include chlorophyll a, chlorophyll c, beta-carotene, luteins and xanthophylls. The pigment fucoxanthin gives it a distinct brown colour
- The growth pattern may be apical (Fucales, Dictyotales), intercalary (Laminariales) or trichothallic (Ectocarpales).
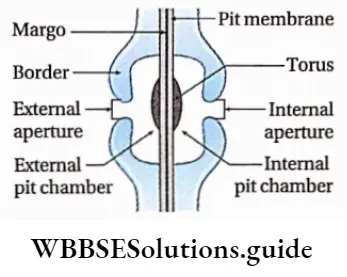
- The cell wall is differentiated into outer mucilaginous and inner cellulose layers. The outer layer contains succinic and alginic acids. The cell wall is covered by a layer of gelatinous hydrocolloid called algin. This protects the cell during unfavourable conditions.
- The cells usually have many small vesicles and white granules. The granules are called fucosan granules.
- Single, stalked pyrenoides are present.
- Motile zoospores and gametes have two laterally inserted unequal flagella. The larger one is tinsel or pantonematic and the smaller one is whiplash or acronematic type.
- The reserve foods are commonly laminarin and mannitol. Sucrose and glycerol are also present in some members.
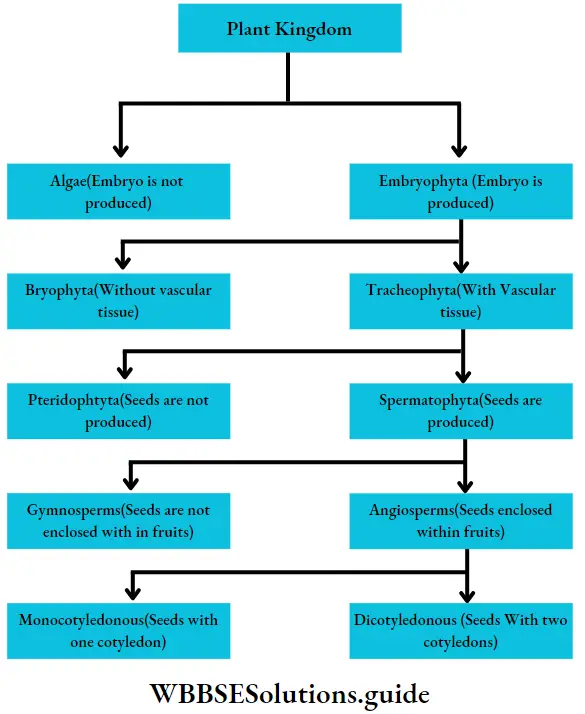
- Reproduction takes place vegetatively (mainly by fragmentation and special propagules), asexually (asymmetrical flagella-containing zoospores or non-flagellate tetraspores) and sexually (isogamy, anisogamy and oogamy).
- In most of the members, fertilisation is external. The zygote does not undergo meiotic division and on germination, it develops into a diploid thallus.
- The life cycle is diplohaplontic.
- The alternation of generations is isomorphic or heteromorphic in nature.
Embryophyta
Embryophyta Definition: The embryophytes are the most familiar sub-kingdom of green plants that form embryos from the zygote after fertilisation.
Embryophyta Definition General features:
- Embryophyta is mainly characterised by the presence of multicellular sporophytes, cuticles, archegonia, antheridia and sporangia, as well as the presence of sporopollenin in spore walls.
- They are a terrestrial species of plants. They produce embryos but do not produce accessory spores.
- The gametangia are covered by a jacket of sterile cells.
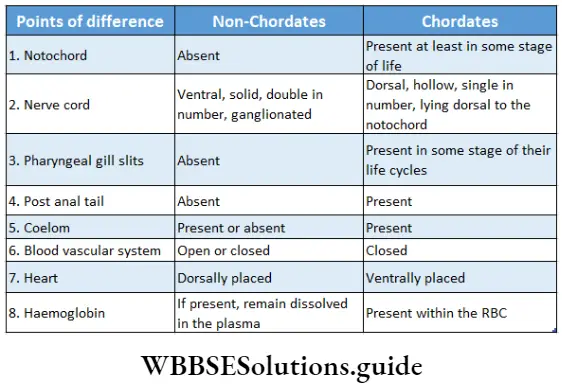
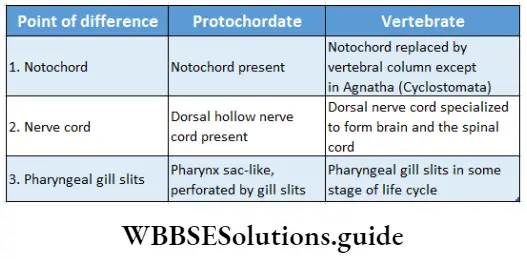
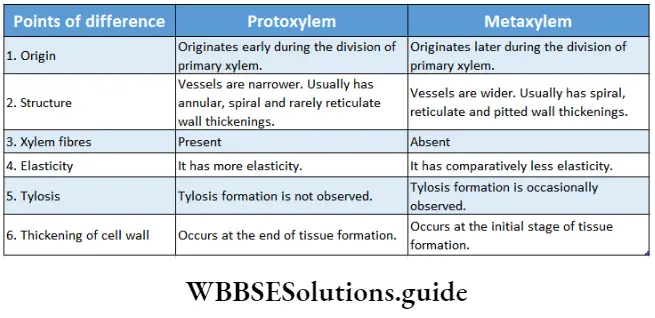
Embryophyta Definition Classification: According to the presence of conducting tissue, embryophyta can be divided into two groups—Bryophyta (without vascular tissue), and Tracheophyta (with vascular tissue). These two groups have been discussed under separate heads.
Tracheophyta Tracheobionta
The term tracheophyta came from two Greek words ‘trachea’ (vascular strand) and ‘phyton’ (plants).
Tracheophyta Definition: Tracheophyta are the major terrestrial plant group, with conducting tissues.
The members of this group are also called vascular plants. Vascular plants include the club mosses, ferns, gymnosperms and angiosperms.
“plant kingdom notes with diagrams and key points”
Tracheophyta General features:
- The tracheophyta is the largest and most advanced group of plants.
- The sporophyte (2n) is the dominant generation in the life cycle of tracheophytes.
- The sporophyte is independent and autotrophic and possesses specialised vascular tissues.
- The plant body is differentiated into true roots, stems and leaves.
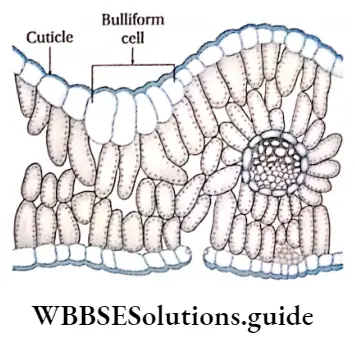
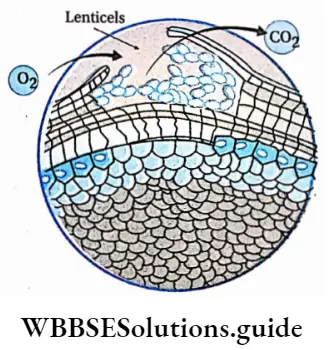
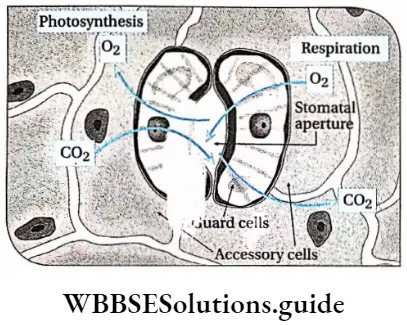
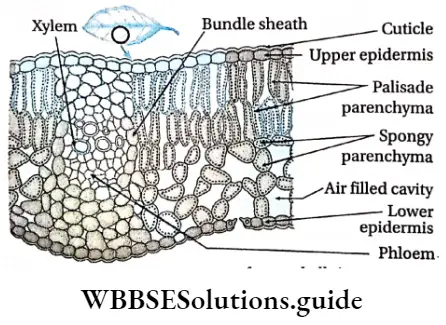
- Leaves are broad, containing a large amount of chlorophyllous tissues. Stomata are present on the leaves, for the exchange of gases.
- Reproduction is mostly vegetative and sexual. Formation of the embryo takes place.
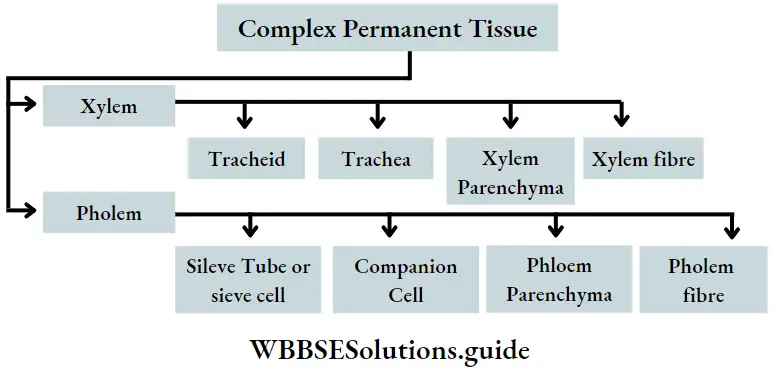
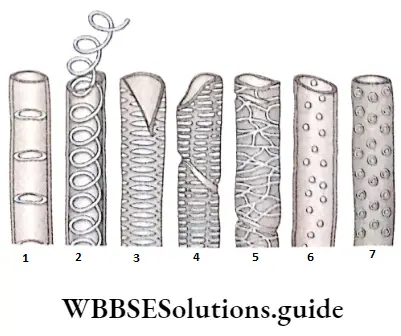
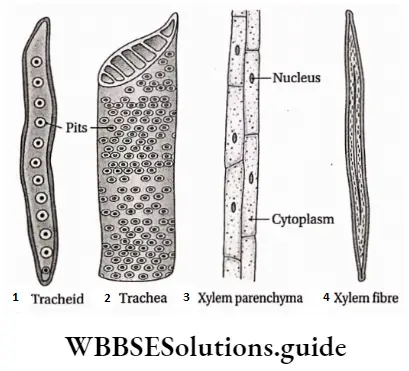
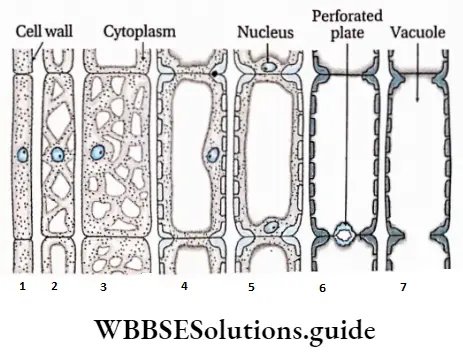
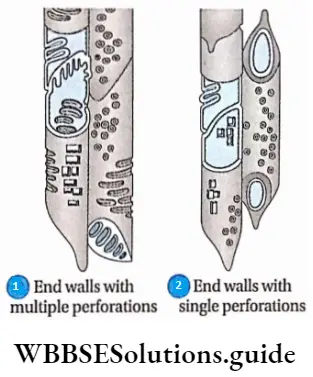
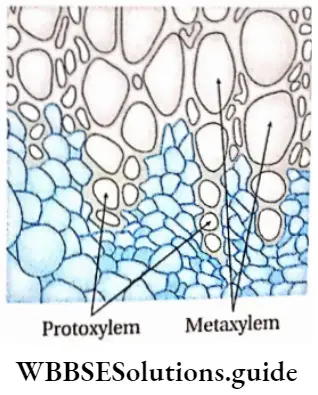
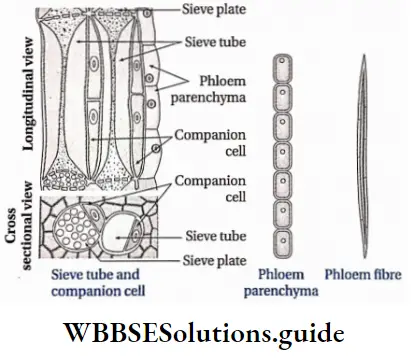
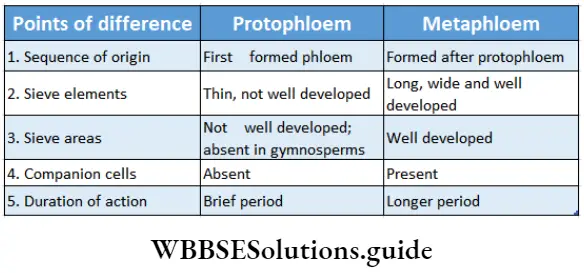
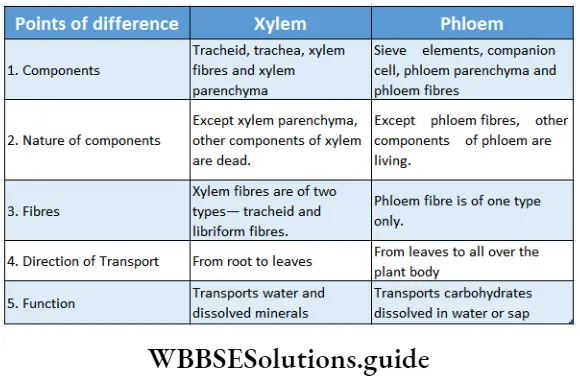
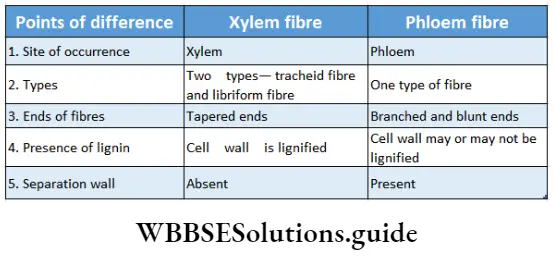
- They contain vascular or conductive tissues—xylem and phloem. The xylem transports water and minerals from soil to leaves. The phloem transfers photosynthetic products from leaves to all other parts of the plant.
- Their size may range from a few millimetres to several hundred metres.
- Their life cycle extends from about some weeks to several years.
Classification of Tracheophyta:
Tracheophytes are classified into two groups—
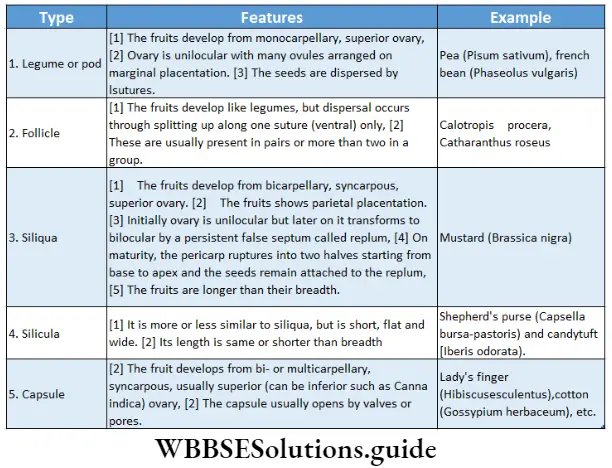
- Pteridophyta (seeds are not produced) and
- Spermatophyta (seeds are produced). These have been discussed below under separate heads.
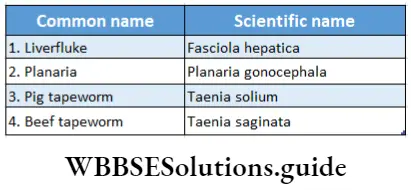
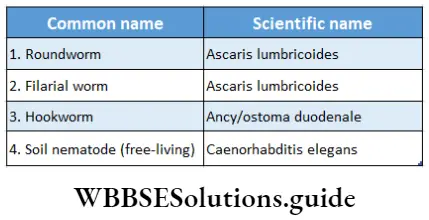
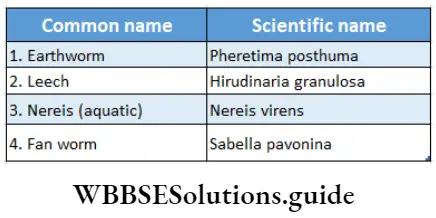
Pteridophyta
Pteridophytes (Greek: pteron-feather, phyton-plants) are non-flowering vascular plants. Hence they are also known as ‘vascular cryptogams’.
Pteridophytes Definition: Pteridophyta is the large group of primitive land plants, with true body differentiation and conducting tissues but does not produce seeds.
They are represented by about 400 genera and about 10,500 species including both living and fossil plants.
Pteridophytes Distribution: Most of the pteridophytes are terrestrial in nature. They are mainly found in damp, shady parts of mountains, damp walls, tree trunks, etc. Aquatic plants like Azolla sp. and parasitic plants like Polypodium sp. are also included in this group.
Pteridophytes Life cycle: Pteridophytes show mainly two types of life cycle—sporophytic and gametophytic.
Characteristics of sporophytes:
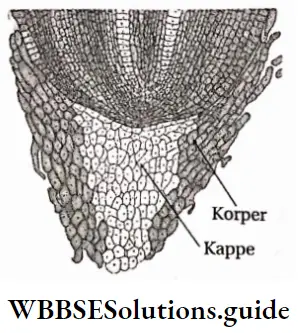
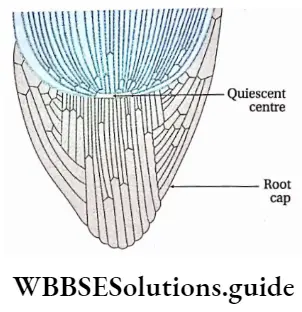
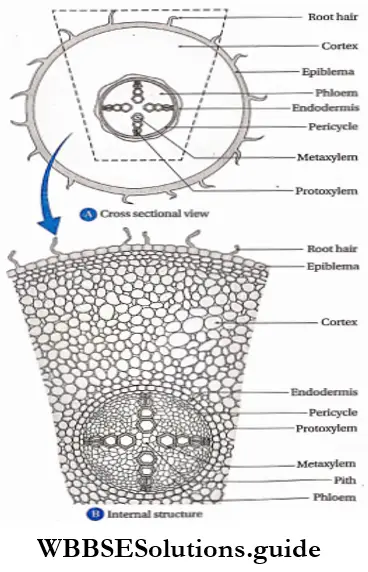
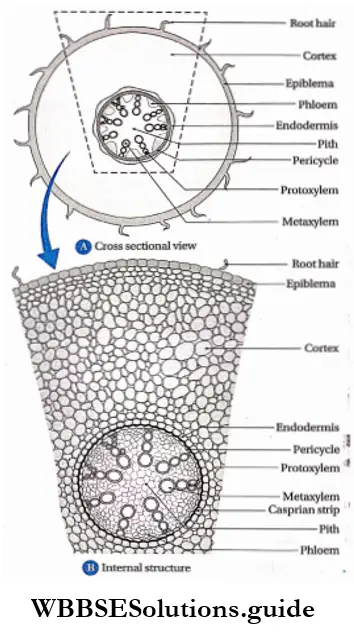
- Pteridophytes are the first true land plants. The predominant generation is the sporophytic plant body, differentiated into true root, stem and leaves. The sporophyte is independent and autotrophic.
- The stem is aerial or rhizomatous. The stem is generally branched—either dichotomous or monopodial. Stem sometimes is densely covered with brown scales.
- The primary root is short-living. The primary root is replaced by a dichotomously branched, adventitious root.
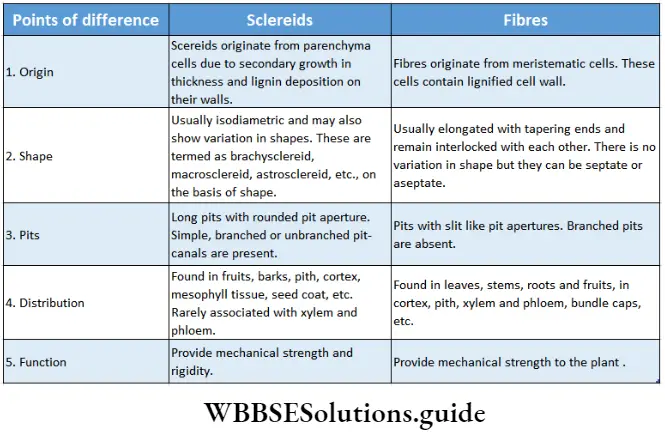
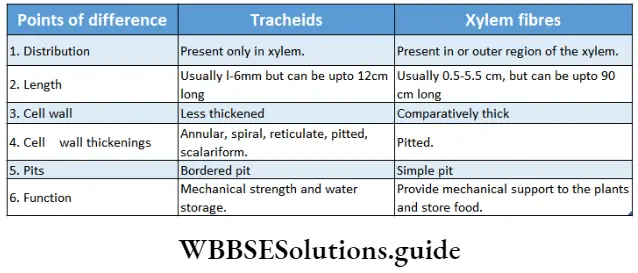
- Conducting tissues (xylem and phloem) are present. But tracheids and companion cells are absent in the xylem and phloem respectively.
- Leaves are of two types— brown-coloured scale leaves and green-coloured sporophylls bearing sporangia, Sporophylls may be small microphyllous and large megaphyllous. The young leaf tips of ferns show circinate ptyxis (arrangement of immature leaf coiled form, which unfolds with maturity).
- Haploid (n) spores are produced within sporangia. In some species (for example Dryopteris) all the spores are similar in structure. They are called homosporous pteridophytes. In some species (for example Marsilea) spores are dimorphic (i.e., larger female spores or megaspores and smaller male spores or microspores). Such plants are known as heterosporous pteridophytes.
- In the case of heterosporous plants, megasporangia are borne on megasporophyll, while microsporangia are borne on microsporophyll.
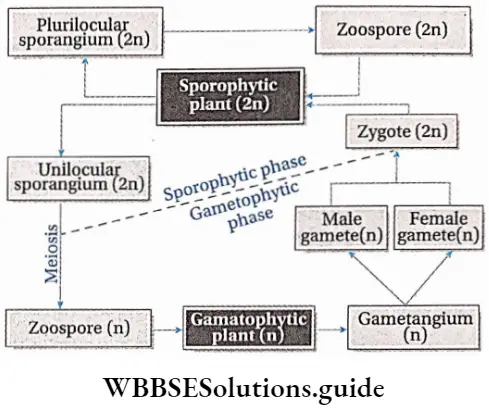
- Some pteridophytes bear a cluster of sporangia called sori (singular: sorus) on either the lower surface of leaves (example Dryopteris sp.) or inside special structures called sporocarps (example Marsilea sp.) or in spike(for example Ophioglossum sp.).
- The sori are covered by a bilayered, smooth membrane, called indusium.
- These spores germinate to form gametophytic plants.
“plant kingdom definition “

Characteristics of gametophytes:
- The gametophytic plant is haploid, autotropic, independently living, dorsiventral, heart-shaped thallus. It is called prothallus.
- Prothalli are rather simple structures and do not have vascular tissues. They may be attached to the substrate by fine multicellular rhizoids. They resemble thallose liverworts.
- In the case of homosporous plants, the spores germinate to produce monoecious prothallus, which produces both antheridia and archegonia.
- In the case of heterosporous plants, antheridia are produced on male prothallus developed from microspores and archegonia are produced on female prothallus developed from megaspores.
- Antheridia are small club-shaped structures, that produce motile sperms or antherozoids. They show chemotactic movement towards archegonia. Archegonia are flusk-shaped structures, with a cup-shaped venter and a long neck. Inside the venter an egg cell is present.
- The zygote is produced after fertilisation and quickly undergoes repeated mitotic divisions to form an embryo. The embryo develops the sporophytic plant.
- Flowers, fruits and seeds are not produced.
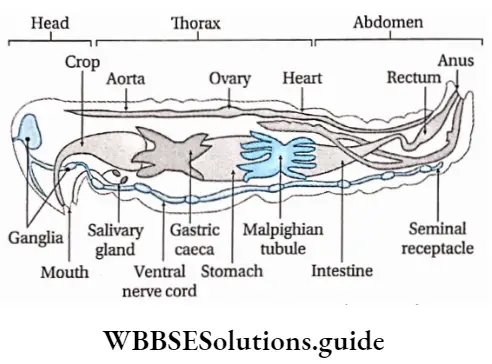
Some common examples of Pteridophytes are— Psilotum sp., Lycopodium sp., Selaginella sp., Isoetes sp., Pellia sp., Equisetum sp., Ophioglossum sp., Pteris sp., etc.
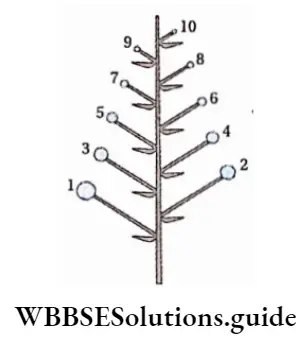
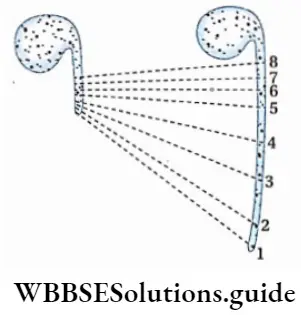
Alternation of generations: All pteridophytes show a diplohaplontic life cycle, heteromorphic alternation of generations.
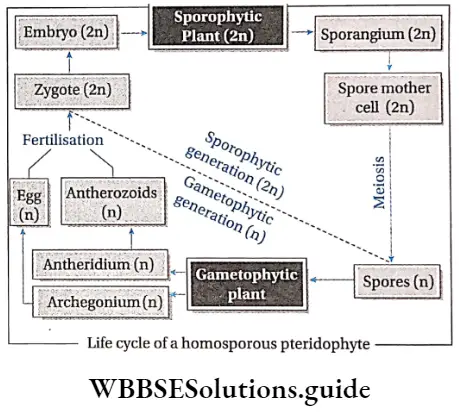
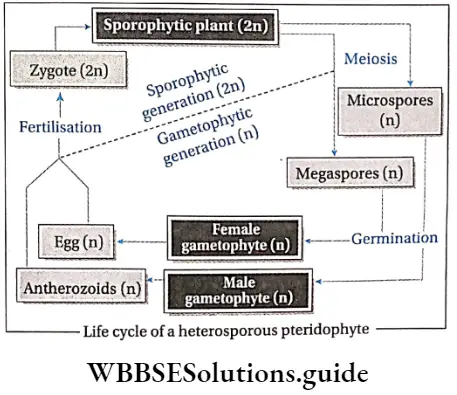
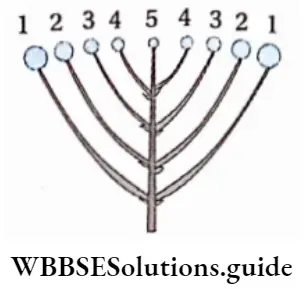
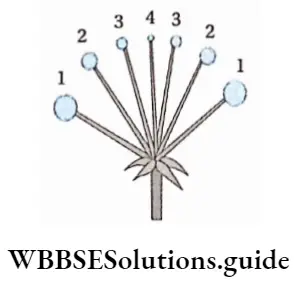
The alternation of generations in the life cycle of a typical pteridophyta (Dryopteris sp.) is shown in the figure below.
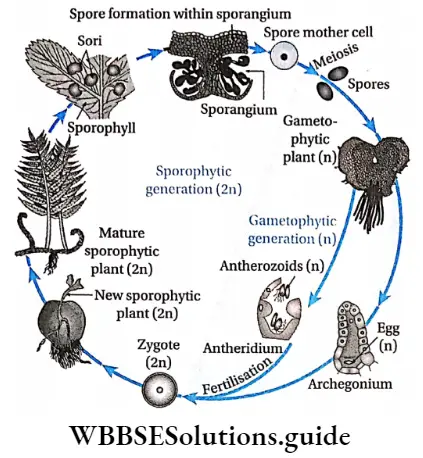
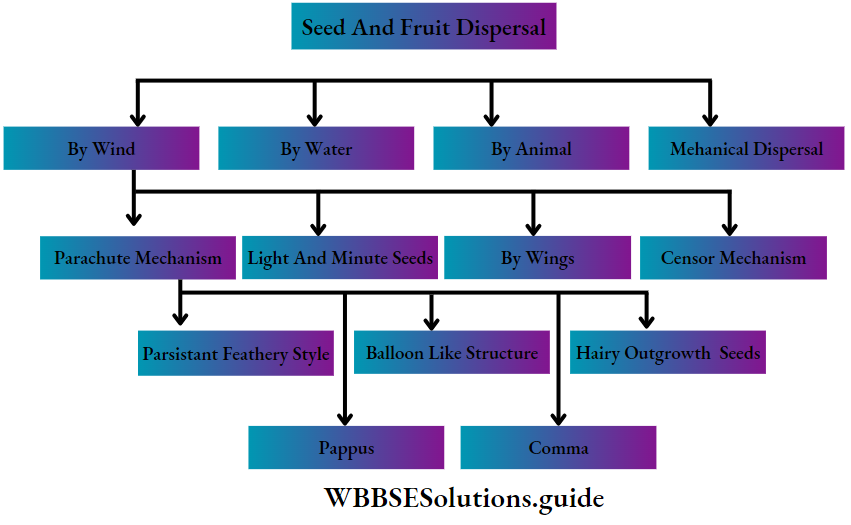
Commercial importance of pteridophyta: Many species of pteridophytes are used for various purposes.
Source of Food: The young leaves or fronds of Ampelopteris proliferate, leaf tips of Matteuccia struthiopteris and leaves of Diplazium esculentum, Marsiiea sp., are also used as vegetables.
Source of fodder: Dry fronds of many ferns are used as the livestock feed for cattle. The leaves of Marsiiea sp. are used as fodder for animals. The rhizome of many ferns such as Pteris sp., which is rich in starch, is used as animal food. The corm (modified stem) of Isoetes sp. is consumed as food by pigs, ducks and other animals.
Source of medicines: Lycopodium sp., Equisetum sp., Selaginella sp., Marsiiea sp., etc., are some pteridophytes that are used to treat neural disorders.
The spores of Lycopodium sp. have been widely used in pharmacies as a protective powder for tender skin and also as water-repellants. The foliages of Lycopodium sp. are used as tincture, powder, ointment and cream and used as a diuretic too.
“plant kingdom classification chart and characteristics”
Equisetum sp. (Horsetail) is rich in silicic acid and silicates. Potassium, aluminium and manganese, along with fifteen types of flavonoid compounds, have been reported from Equisetum.
The flavonoids and saponins are assumed to cause the diuretic effect. The silicon is believed to exert connective tissue-strengthening and anti-arthritic action.
Several ferns are used as herbal medicine. Oil extracted from Aspidium rhizome is used as a vermifuge (medicine against worms). The leaf extract of Polypodium is used to cure stomach and liver problems.
The root extract of Osmunda regatis is used for the treatment of jaundice. The root extract of Osmanda vulgaris is used to heal wounds.
Aesthetic uses: Many species of pteridophytes are cultivated for their aesthetic value. Some epiphytic species of Lycopodium sp. are grown on hanging baskets.
Several species of Selaginella are used in decoration during festive occasions due to their decent foliage and colour. Several ferns such as Angiopteris sp., Aspienium sp., Marattia sp., etc., have beautiful soral arrangements, hence used for ornamentation.
Polishing purpose: Accumulation of silica in the stem cells of Equisetum sp. gives it rough structures. Thus, it can be used as a scrubber for polishing purposes.
Preparation of fire-crackers: Since the spores of Lycopodium sp., are highly inflammable, hence they can be used to prepare fire-crackers.
Biofertilisers: Azolla sp., is a free-floating water fern which can grow very quickly through vegetative propagation. They live symbiotically with a nitrogen-fixing cyanobacteria—Anabaena sp., etc.
The alga provides nitrogen to the plant and to the growing aquatic medium. Thus, Azolla sp., growing in rice fields serves as a green manure for better crop yield.
Metal indicators: Equisetum sp., accumulates minerals, especially gold, in their stem. The rate of accumulation even reaches up to about 4.5 ounces per ton.
They may be referred to as gold indicator plants, which help in identifying a region with gold ore deposits. Similarly, Aspienium adulterinum is an indicator of nickel and Actinopteris australis is a cobalt indicator plant.
Fossil fuels (coal): During the carboniferous period, the flora of the earth was dominated by huge pteridophytes (Lycopods), ferns and other large leafy plants. The coal formed after their death had become sources of different fossil fuels.
Classification of pteridophyta: Pteridophytes have been classified into four major classes by Doyle (1971)—Psilopsida, Lycopsida, Sphenopsida and Pteropsida.
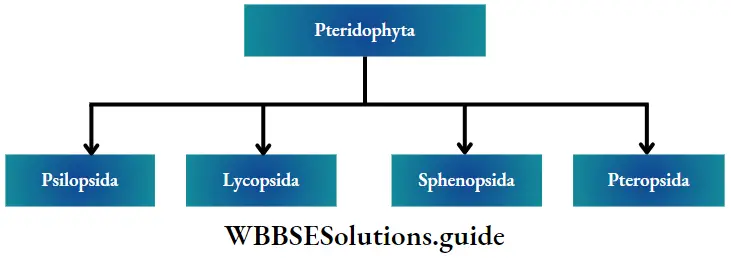
Fossil fuels General features:
- They are mainly terrestrial pteridophytes, found in warm climates.
- The plant body is a rootless sporophyte that differentiates into a subterranean (just below the earth’s surface) rhizome and an aerial erect shoot.
- Branching is dichotomous in both rhizome and shoot.
- The large rhizoids borne on the rhizome, absorb water and nutrients from the soil.
- The aerial shoots bear spirally arranged scale-like (for example Psilotum sp.) or leaf-like appendages (for example Tmesipteris sp.).
- Stele is protostelic or siphonostelic with sclerenchymatous pith.
- Cambium is absent Hence, secondary growth is absent.
- Bi or trilocular sporangia are borne in the axils of leaf-like appendages.
- The mode of sporangial development is of eusporangiate (more than one cell initiates sporangial development)type.
- They are homosporous.
- The gametophytes or prothalli are independent, colourless, cylindrical, branched and subterranean. They grow as saprophytes with an associated endophytic fungus.
- Antherozoids are spirally coiled and multi-flagellated. Examples: Psilotum sp. (living); fossil members are- Rhynia sp., Cooksonia sp., Zosterophyllum sp etc.
Lycopsida (Lycopods)
Lycopsida General features:
- Thin and small structured pteridophyta, found mostly in forests and mountains.
- The sporophytic plant body is differentiated into definite root, stem and leaves.
- The sporophytes are dichotomously branched.
- The leaves are usually small and microphyllous. The leaves are either isophyllous or dimorphic. In some species, ligules (tongue-shaped membranous structures) may present towards the leaf apex.
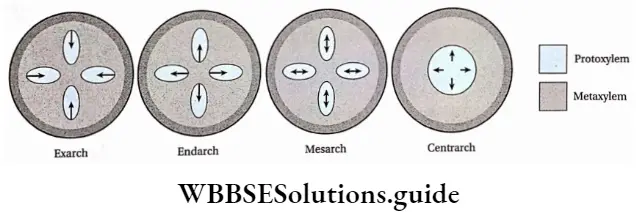
- The xylem arrangement in the stem is exarch.
- Stele may be protostele, siphonostele or polystele in nature
- Sporangia are borne singly on the adaxial (upper) surface of the sporophylls. In most cases, sporophylls join to form cones or strobili.
- The spores are homosporous (for example Lycopodium sp.) or heterosporous (for example Selaginella sp.).
- The spores develop into independent gametophytes.
- Antherozoids are flagellated.
Examples: Lycopodium sp. and Selaginella sp. are living genera and there are 14 extinct genera—Asteroxylon sp., Baragwanathia sp., etc.
Sphenopsida (Horsetails)
Sphenopsida General features:
- The living members of this group are found in damp and shady regions of mountains, forests and marshlands.
- The stems and branches are differentiated into nodes and internodes. Internodes are with longitudinal ridges and furrows.
- Branches arise in whorls.
- The leaves are extremely reduced and borne in whorls at the nodes of aerial branches and stems.
- Stele is Protostele or siphonostele in nature.
- The sporangia develop on an elliptical pallet appendage called sporangiophore. Sprorangiophores are arranged into a cone shaPed strobilus (P|ural: strobila)-
- Most of the members are homosporous including Equisetum sp. However, some extinct forms were heterosporous (for example Catamites Hashana).
- The gametophytes are exosporic (develop outside the spore) and green.
- Antherozoids are multiflagellate.
Examples: This class is represented by a single living genus and 18 extinct genera. The living genus is Equisetum sp. and the fossil genera are Catamites sp. Annularia sp., etc.
Pteropsida
This group is commonly known as ferns. The pteropsida differs from other classes in having large leaves (fronds). This is the largest and highly evolved group of pteridophytes and is represented by about 9,000 species which show a wide range of distribution. Pteropsida are known from as far back as the Devonian period of the Paleozoic Era.
Pteropsida General features:
- Members of Pteropsida are found in shady and damp forests, high altitudes of mountains, marshlands and moist soil of plains. Some members are aquatic and some are epiphytic.
- The sporophytes are usually perennial in nature and differentiated into roots, stems and spirally arranged leaves.
- The stem is rhizomatous, mostly short and stout. Adventitious roots grow from it.
- The rhizome is covered with brown scales.
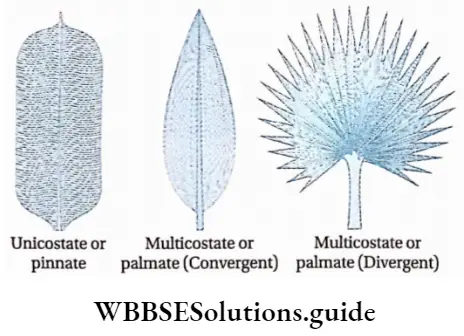
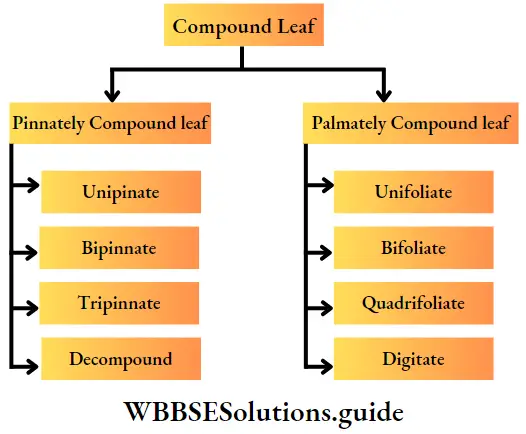
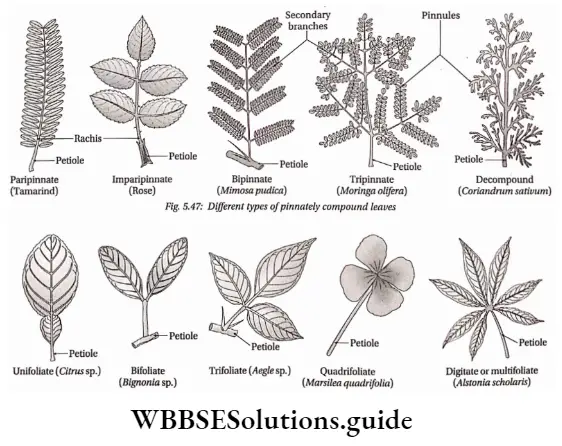
- The leaves are large, simple (Ophioglossum sp.) or pinnately compound (Dryopteris sp.) and are called fronds. In Adiantum sp., the rachis is dichotomously branched and bears a fan-shaped leaflet.
- The rachis is covered with tiny brown hairs called ramenta.
- Young fronds show deracinate ptyxis, except Ophioglossum sp.
- The stele can be of different types, for example protostele, siphonostele, etc.
- Vegetative reproduction fragmentation, adventitious buds and bulbils (fleshy globose structures). In Adiantum sp., new plants, are produced from leaf tips when they come in contact with the soil.
- Matured vegetative leaves bear sori on their lower surface. They are then known as sporophylls. Sori bear a number of sporangia. Sporangial clusters are sometimes enclosed by a shield-like structure called indusium.
- These plants are homosporous in nature. Haploid spores are formed through meiosis from diploid spore mother cells.
- Monoecious gametophytes or prothalli are produced by the germination of spores. Rhizoids present on prothallus.
- Antherozoids are ciliated and coiled.
Examples: Ophioglossum sp., Pteris sp., Dryopteris sp., etc.
Similarities between bryophyte and Pteridophyta
- Both are non-flowering and terrestrial.
- Both have multicellular and multiflagellate reproductive organs.
- Both their antheridia and archegonia are similar in structure, with a layer of sterile cells covering them. Antheridia are club-shaped and archegonia are flask-shaped in both.
- Antherozoids are coiled and motile in both.
- Both have a definite alternation of generations.
- Asexual reproduction takes place by spores.
Spermatophyta
Spermatophyta Definition: Spermatophyta is the higher group of flowering land plants where seeds are produced and embryo development takes place.
Spermatophyta Distribution: They are found all over the land and sometimes in water as well.
Spermatophyta General features:
- The body is differentiated into roots, stems and leaves. Vascular tissues (xylem and phloem) and mechanical tissues, both are present.
- The plants are heterosporous with microspores and megaspores.
- Gametophytic plants are dependent on sporophytic plants.
- The reproductive organs are multicellular. Instead of being released, the megaspores remain within the ovules where one or more female gametes are formed.
Classification of the Spermatophyta: The spermatophytes are further classified into two groups
These topics are discussed under separate heads.
Cycadopsida
Members of the class Cycadopsida produced the most dominating flora during the mid-Mesozoic Era.
Cycadopsida General features:
The general features of Cycadopsida are as follows—
- Plants generally with stout trunks having manoxylic wood. They have large pith regions but have woody cortexes.
- Leaves are large and frond-like. They are basically pinnately compound in form or in venation.
- Annual rings are absent.
- Presence of ciliated sperms.
- These plants are usually dioecious with the reproductive organs borne on specialised leaves (sporophylls).
- The pollen grains (microspores) form within the microsporangia on the sporophylls of the male cones.
- The ovules (megaspores) develop without protective coverings on the sporophylls of the female cones.
- The pollination is carried out by wind or beetles.
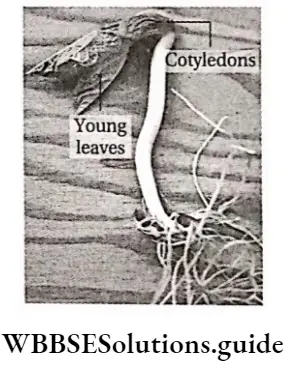
- After fertilisation, the seed develops without a protective pericarp. The seed is radially symmetrical. The seed has an outer fleshy layer and the embryo has two cotyledons.
- The life cycle is diplohaplontic, alternation of generations is heteromorphic. The life cycle of cycads has two distinct phases. The gametophyte stage is microscopic and enclosed within the microspores and megaspores.
Examples: Cycas sp., Ceratozamia sp., Zamia, etc.
Ginkgopsida
The class Ginkgopsida includes a single order of Ginkgoales—now represented by a single living member Ginkgo biloba. The members of this group first appeared in the Permian period, but gradually depleted from the earth.
The leaf impressions of G. biloba have been identified from the Permian rocks and probably from the Carboniferous rocks. G. biloba has escaped extinction and still exists today being the oldest living seed plant Thus, Ginkgo biloba is referred to as a ‘living fossil’.
Ginkgopsida General features:
The general features of Ginkgopsida are as follows—
- The members of this group are mostly tall, woody, deciduous and branched trees. They have both long and dwarf shoots.
- Secondary wood is pyroxylin i.e., has a large number of thick-walled tracheids and fibres.
- Leaves are leathery, strap-shaped or fan-shaped, often with dichotomous venation.
- These plants are dioecious, i.e., both male and female gametophytes are present on the same plant. Produce both male and female cones.
- Male fructifications are axillary, unbranched catkin-like bearing microsporangiophores.
- Each microsporangiophore bears 2-12 pendulous microsporangia.
- Ciliated sperms are present.
- Ovules are 2-10 in number, terminal on axillary branching or almost unbranched axes.
- Seeds are large showing radial symmetry.
- Seeds have with fleshy outer layer and a stony middle layer.
Examples: Ginkgo biloba
Coniferopsida
The Coniferopsida includes four orders—Cordaitales, Voltziales, Coniferales and Taxales. They probably appeared during the Upper Devonian and were the most dominating taxa during the Upper Carboniferous to Triassic. At present, this class is represented by 57 living genera.
Coniferopsida General features:
The general features of Coniferopsida are as follows—
- Mostly tall, woody, evergreen plants which are profusely branched.
- Secondary wood is pyroxylin.
- They have needle-shaped, paddle-shaped or fan-shaped foliage leaves. The arrangement of leaves is whorled or a|ternate. The )eaves are covered with a thick cuticle.
- Long and dwarf shoots are present.
- Stele in the stem are endarch, pith and cortex are small, with large conducting tissues. The woody part is more in these plants.
- Resin canals are often present in leaves and stems.
- The plants are usually dioecious. The reproductive organs are borne on specialised leaves (sporophylls), which are compactly arranged in cones or strobili.
- Male cones or microcstrobili are smaller and survive for less number of days. Female cones or megastrobili are large, and long and survive for longer periods of time.
- The gametophyte stage is represented by microspores or pollen.
- Male gametes are represented by the male nucleus only.
- The pollen forms within the microsporangia present on the scales (microsporophylls) of the male cones. They are often winged.
- The ovules are borne on the sporophylls of the female cones and the pollen is usually transferred to the ovules by wind.
- After fertilisation, the seed develops directly on the female sporophylls.
- Seeds are bilaterally symmetrical. The embryo has two too many cotyledons.
Examples: Pinus sp., Picea sp., Abies sp., Cedrus sp., etc.
Gnetopsida
The class Gnetopsida is considered to be the highly evolved group of gymnosperms. The class comprises three distinct orders—Ephedrales, Welwitschiales and Gnetales, with a monogeneric family in each. However, some scientists prefer to retain them within a single order, Gnetales, with a monogeneric family in each.
Gnetopsida General features:
The general features of Coniferopsida are as follows—
- The members of this group are small trees or shrubs or lianes in nature.
- The plants bear opposite or whorled leaves, with reticulate venation.
- Vessels are found in the secondary xylem. For that reason, they are considered to be ancestral to angiosperms.
- Resin canals are absent.
- Flowers are monoecious. Male and female cones are borne separately.
- Compound male and female strobili resemble the inflorescence of angiosperms.
- Male flowers consist of a stalk bearing two or more sporangia. They may bear bract and perianth.
- Female flowers consist of ovules only.
- They show unique fertilisation features i.e., a pollen tube grows in order to fertilize the egg. Thus, the sperms themselves are not motile, like other gymnosperms.
- The embryo contains two cotyledons.
Examples: Gnetum ula, Ephedra sp., Welwitschia sp., etc

Similarities between Pteridophyta and gymnosperms
- Both have a major sporophytic phase and a less important gametophytic phase.
- Both have sporophytic plant bodies, differentiated into roots, stems and leaves.
- Both have vascular tissues, without vessels in the xylem (exception: Gnetopsida) and companion cells in the phloem.
- Pteridophytes are heterosporous and have motile male gametes (reproductive cells) like many gymnosperms (for example Cycas sp.).
- Both show definite alternation of generation.

Angiosperms
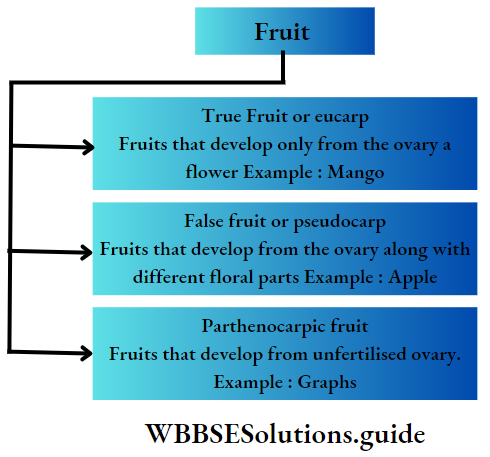
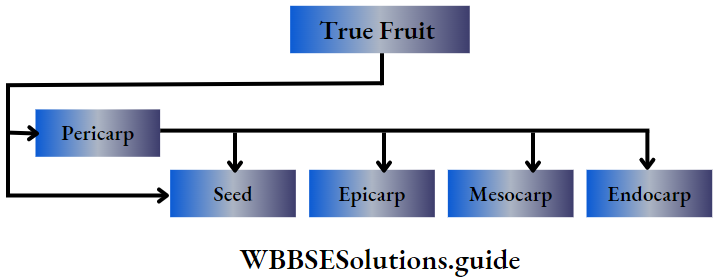
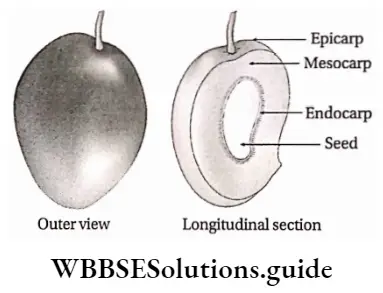
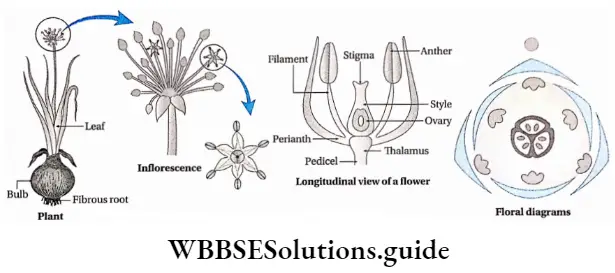
Angiosperms Definition: Angiosperms are flowering plants that produce fruits, that enclose the seeds.
Angiosperms are the highest-order plants in the plant kingdom. They are the largest group of plants, that are flowering and contain seeds in special structures, called fruits.
Angiosperms Distribution: Angiosperms originated quite late on earth, yet they occupy the majority of it. Most of them are terrestrial, and only a few are aquatic. They are found under the ocean, as well as at 6000m above sea level.
Angiosperms General features:
- The main plant body is sporophytic in nature. The sporophytic stage is independent and diploid (2n). The gametophytic phase is haploid (n) and dependent on the sporophytic phase.
- Angiosperms are trees, shrubs or herbs in nature. The trees and shrubs are generally perennials, while the herbs are annuals, biennials or perennials.
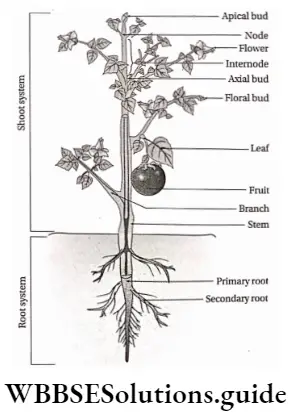
- An angiospermic plant consists of different organs—root, stem, branches, leaves, flowers, fruits and seeds.
- The body of an angiosperm plant consists of two systems— the root system and the shoot system.
- The roots may be taproot or adventitious.
- The stem may be branched or unbranched. In some cases, they may be climbers or lianes.
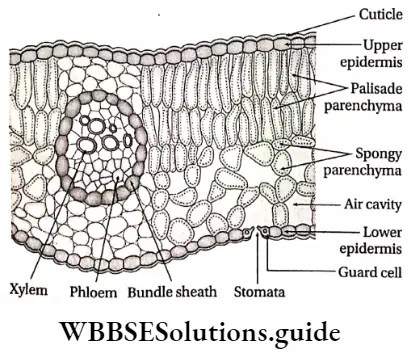
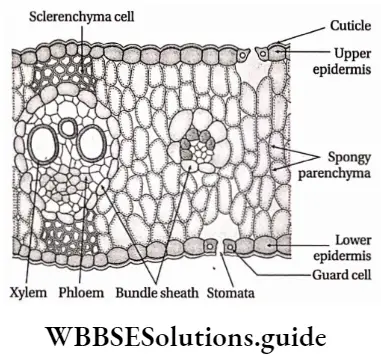
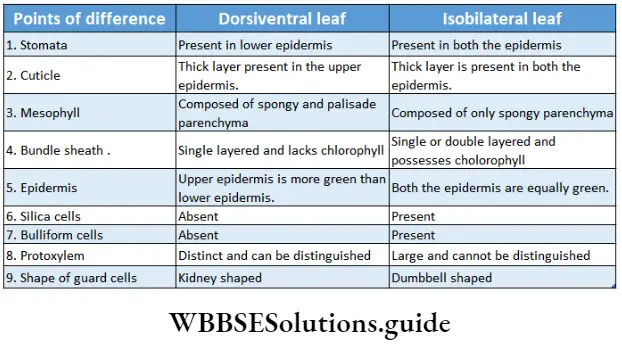
- The leaves are simple or compound, dorsiventral or isobilateral.
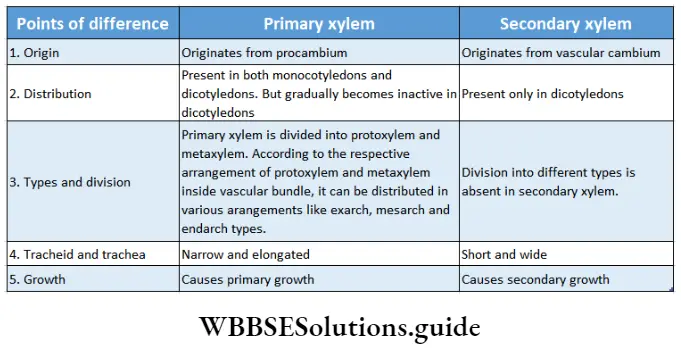
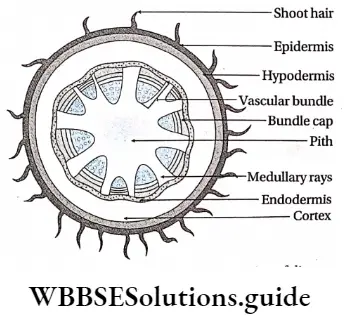
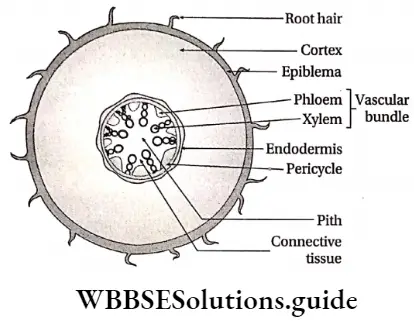
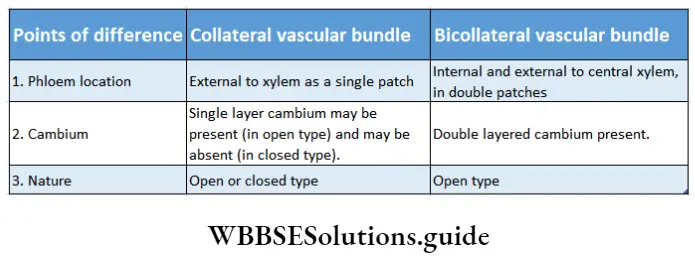
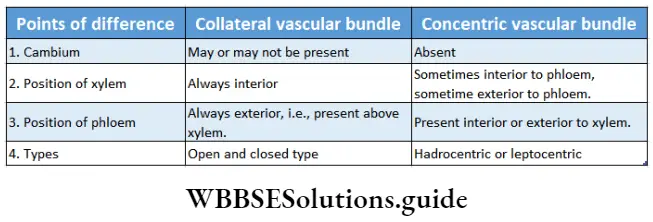
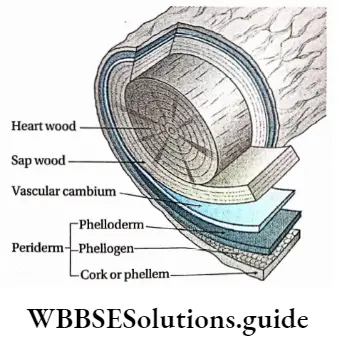
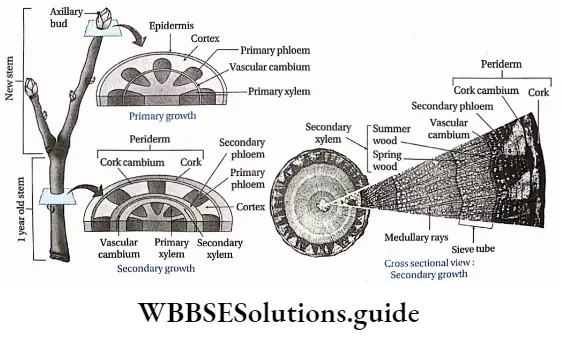
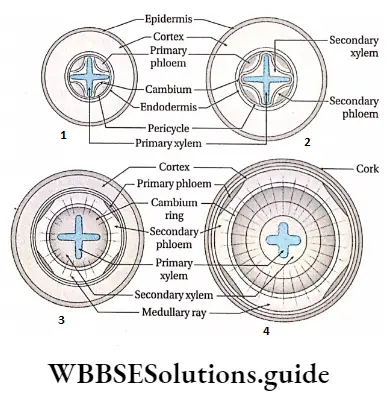
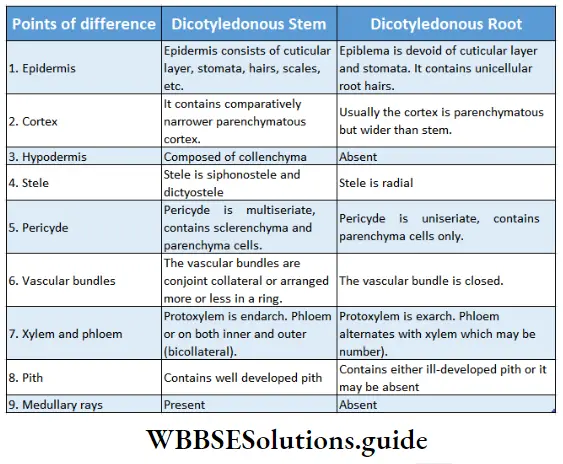
- The conducting tissues are the xylem and phloem. The vascular bundle of roots are radial and exarch, while that of the stem is collateral or collateral, free or enclosed and endarch. Secondary growth is observed in dicots.
- Angiospermic plants contain chlorophyll in their leaves and thus are able to photosynthesise.
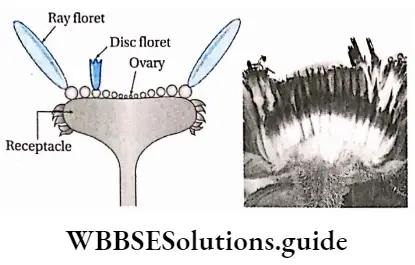
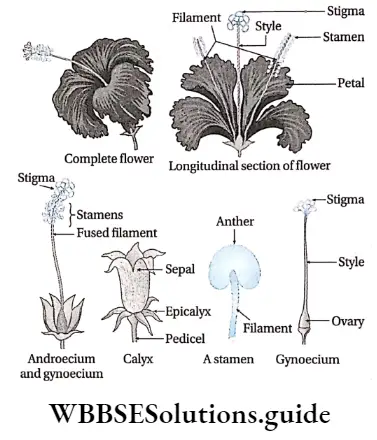
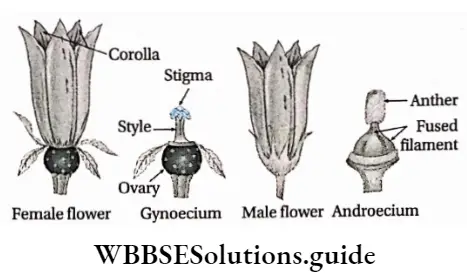
- Flowers may be complete (all four floral whorls are present), incomplete, bisexual, unisexual (male or female), or sterile in nature. They have accessory whorls (sepals and petals), along with reproductive whorls (androecium and gynoecium).
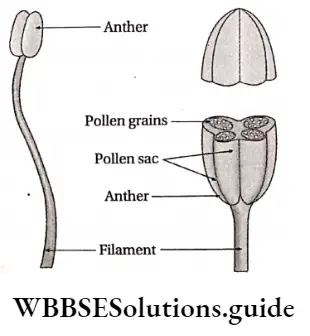
- Androecium has an anther and filament. The anther produces pollen grains.
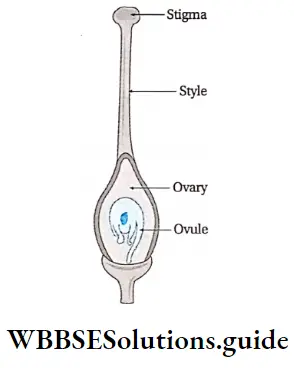
- The gynoecium has an ovary, style and stigma. One or more ovules are produced within the ovary.
- The male gametophytes form pollen tubes. They contain two nuclei.
- The archegonia are not produced within female gametophytes. Instead, the female gametophytes get reduced to four megaspores.
- Only one of the megaspores develops into an embryo sac which contains an egg nucleus. Double fertilisation, occurs in an egg angiosperms. Apart from a zygote, a triploid endosperm is also produced from this.
- After fertilisation, the ovule transforms into seed and the ovary into fruit.
- The zygote develops into an embryo by repeated cell division. After germination, it grows into an adult plant.
Sexual Reproduction in Flowering Plants: Various steps of the sexual reproductive procedure of flowering plants are discussed below.
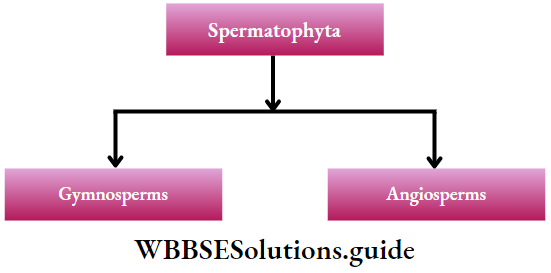
Microsporogenesis: Microspore i.e., the pollen grain is the first cell of the male gametophyte, which contains only one haploid (n) nucleus. During the early stage of development, four microspores or pollens (n) are formed from each microsporocyte or pollen mother cell through meiosis.
Pollination: Transfer of pollen grains to the stigma takes place by several agents, such as wind, water, insects, etc. This process is called pollination. It takes place by two means—self-pollination (within the same flower or two different flowers of the same plant) and cross-pollination (between two different flowers of two plants).
“short notes on plant kingdom for NEET and board exams”
Development of male gametophyte: Firstly, the pollen cell (n) undergoes mitotic division and forms a small generative cell and a large vegetative or tube cell. The generative cell further divides and gives rise to two male gametes. This is known as the 3-celled stage containing a vegetative cell and two male gametes.
The division of the generative cell may take place either in the pollen grain or in the pollen tube. The pollen tube comes out through germ pores after pollination. The nucleus of the vegetative cell, commonly known as the tube nucleus, usually degenerates with the maturation of the generative cell.
Development of megaspore mother cell: The ovule, inside the ovary, develops as a multicellular placental outgrowth including the epidermal and numerous hypodermal cells. This structure develops into a mass of tissue called nucellus, with one or two integuments.
One of the hypodermal (just below the outermost layer) cells of the nucellus enlarges in size and has a dense cytoplasm and a conspicuous nucleus. This is called an archesporial cell, which subsequently divides transversely and forms an inner primary sporogenous cell and an outer primary parietal cell.
The inner primary sporogenous cell functions as a megasporocyte or megaspore mother cell and the primary parietal cell through repeated vertical divisions forms layers of parietal cells. Sometimes, the archesporial cell does not divide and directly functions as a megaspore mother cell.
Megasporogenesis: The diploid megaspore mother cell meiotically divides to form four haploid (n) megaspores, within the ovule. Two subsequent transverse divisions form four, linearly arranged, haploid megaspores (linear tetrad).
Out of four megaspores, only one moves towards the chalazal end (mostly towards the stalk) and becomes the functional megaspore.
The other three, remain at the micropylar end (the small opening of the integument), where they gradually disintegrate. The functional megaspore enlarges to form the female gametophyte i.e., the embryo sac.
Development of female gametophyte: Megaspore (n) is the first cell of the female gametophyte. The functional megaspore enlarges and forms the embryo sac. The nucleus of this embryo sac through three successive divisions forms eight nuclei.
Only one of these eight nuclei forms the egg cell (n). Four of these eight nuclei reside towards the micropylar end and the remaining four towards the chalazal end. A single nucleus from each pole then moves towards the centre and forms a pair of polar nuclei, which then fuse to form a diploid nucleus, called the definitive nucleus (2n).
This is also called a polar fusion nucleus or secondary nucleus. The three nuclei at the micropylar end (one egg cell and two synergids) form the egg apparatus and the rest three at the chalazal end, are called antipodal cells.
In the egg apparatus, each nucleus remains surrounded by a viscous mass of cytoplasm without any wall. The m‘dd*e largest one is called egg or ovum or oosphere and the rest two on each side of the egg are the synergids. The antipodal cells have a viscous mass of cytoplasm, each covered by a cellulosic wall.
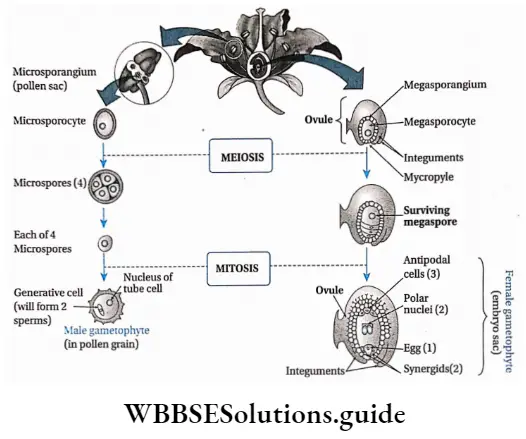
Fertilisation: Fertilisation is a process where male and female gametes fuse with each other to form a zygote, which later on forms the multicellular embryo. In angiosperms, the embryo sac remains embedded into the ovarian cavity.
“types of plants in plant kingdom and their characteristics”
The pollen grains reach the stigma through pollination. The pollen grain germinates on the stigma and forms a pollen tube which grows through the style and reaches the ovule. Here it releases the male gametes near the egg.
One of these male gametes fuses with the egg, to form the diploid zygote (2n). The other male gamete fuses with the definitive nucleus (2n) to form the endosperm nucleus (3n). This gives it the name, double fertilisation.
Germination of pollen grains
The pollen grain usually develops a single pollen tube (monosiphonous), but occasionally in some cases, it may; develop more than one tube (polysiphonous). Sometimes the single pollen tube may be branched.
The commercial importance of angiosperms:
Human civilisation is dependent on plants. The three major necessities of human beings— food, clothing and shelter, are fulfilled mainly by plants.
Food source:
Plants yield various types of food such as—
- Cereals, i.e., rice, wheat, maize, etc.
- Millets, i.e., small-grained food such example, jowar, bajra, ragi, etc.
- Pulses, like, gram, pea, pigeon pea, lentil, mung, etc.
- Vegetables, like, radish, carrots, sweet potatoes, etc.
- Fruits, like, orange, banana, apple, guava, mango, etc.
Food adjuncts: Plants yield spices, flavouring materials, beverages, etc.
- Spices and other flavouring materials like ginger, turmeric, cinnamomum, cloves saffron, pepper, etc.
- Beverages like tea, coffee, etc.
Production of medicines and drugs:
- Medicinal plants such as Cinchona sp., Ipecac sp„ Rauvolfia sp. etc., are sources of various alkaloids. These alkaloids are used to produce medicines.
- Many angiosperms are sources of fumitories and masticatories, for example, tobacco, °P‘um’ marijuana (ganja), etc.
Industrial use of plants and plant products: Plants yield fibres (for example jute, cotton, hemp, etc.), timber (for example teak, sal etc.), and essential oils (for example Eucalyptus, Citronella, etc. sugars (for example sugarcane, palm), cellulose products etc. Each of these has different uses in our daily lives.
Fuel: Dead remains of plants and woody plant parts are used as fuel.
Similarities between gymnosperms and angiosperms
- Both have major sporophytic phases. The gametophytic phase was dependent on the sporophytic phase.
- Both have their body differentiated into roots, stem and leaves. Both produce flowers.
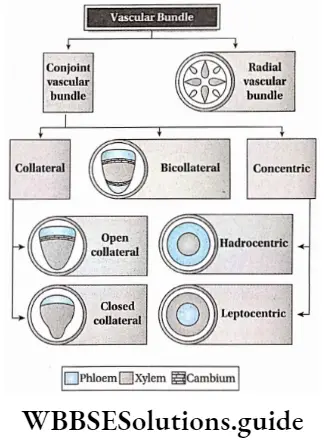
- Both have open or closed collateral vascular bundles.
- Both show the growth of pollen tubes.
- Both are heterosporous plants in nature.
- In both of them, female spores remain within sporangia or ovules.
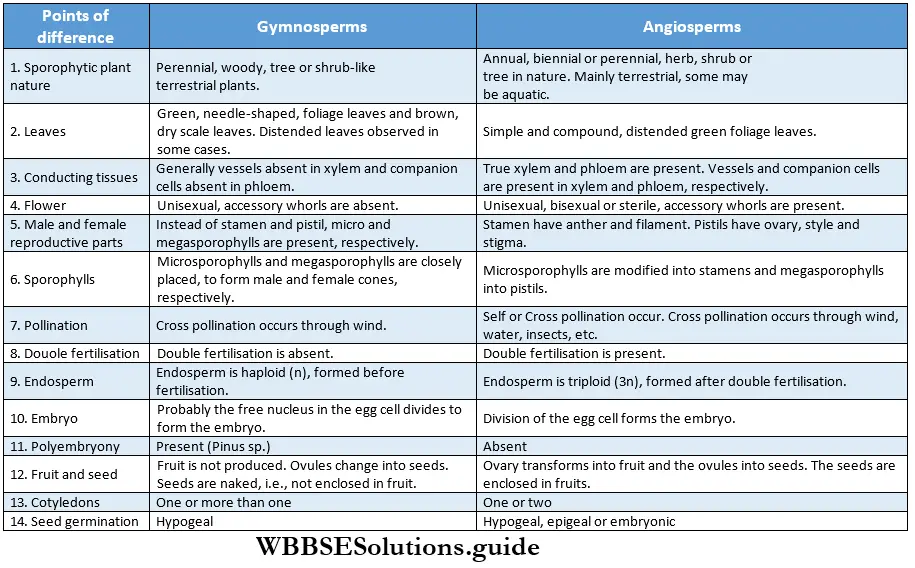
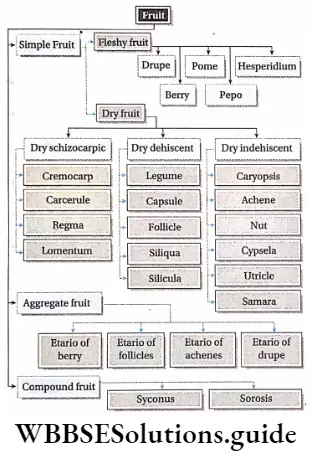
Classification Of Angiosperms
Bentham and Hooker’s System: Bentham and Hooker worked together for about 25 years and jointly published their work in the book Genera Plantarum (1862-1883) in three volumes.
They divided the Angiospermae into two major classes—
- Monocotyledonae (comprises plants having single cotyledon in seed, i.e., monocotyledonous plants) and it was divided into seven series. The Series were further divided into cohorts (= orders), and cohorts into natural orders (= families). Natural orders include genera and finally, each genus includes species.
- Dicotyledonae (comprises plants having two cotyledons in seed, i.e., dicotyledonous plants). It was further divided into three sub-classes—Polypetalae, and Monochlamydeae. Polypetalae was divided into three series; Gamopetalae was divided into three series and Monochlamydeae into eight series. These groups are discussed under separate heads.
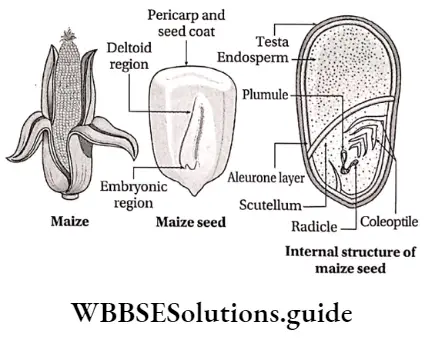
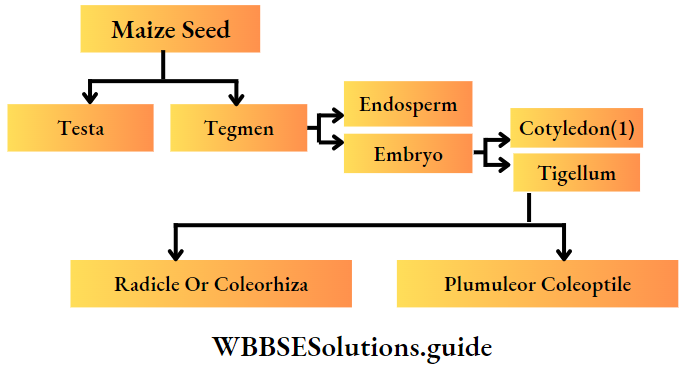
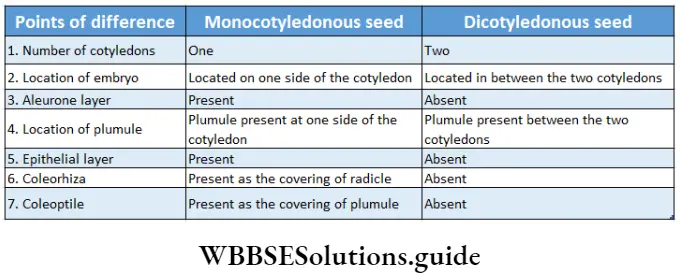
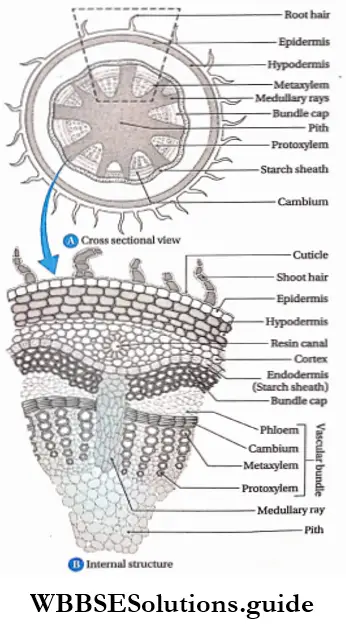
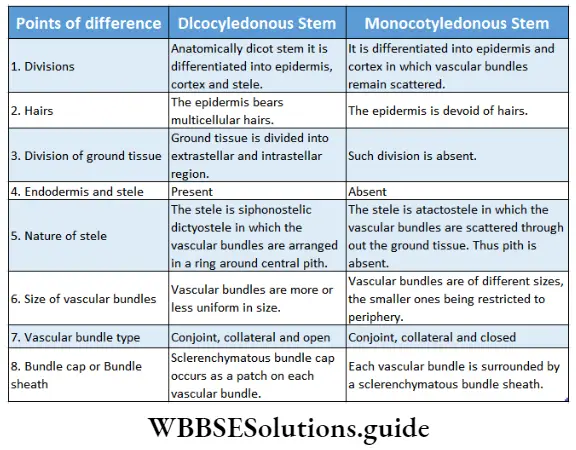
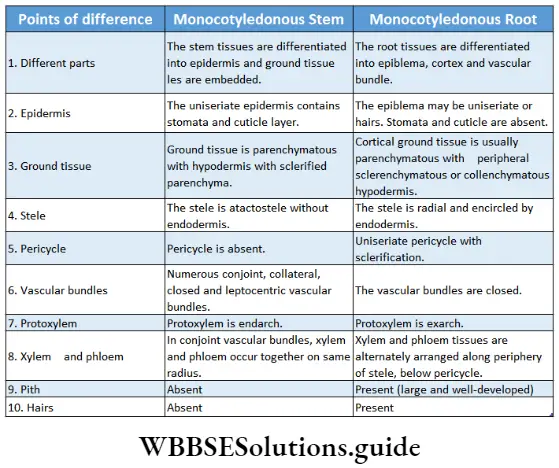
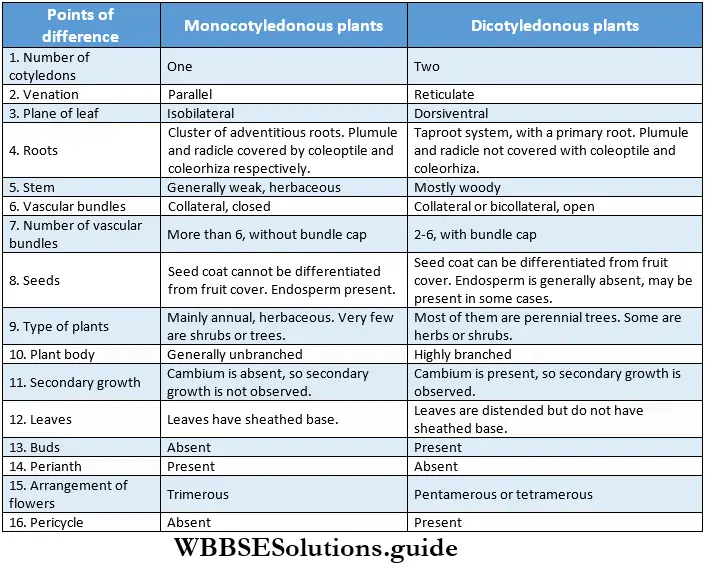
Monocotyledonous Plants
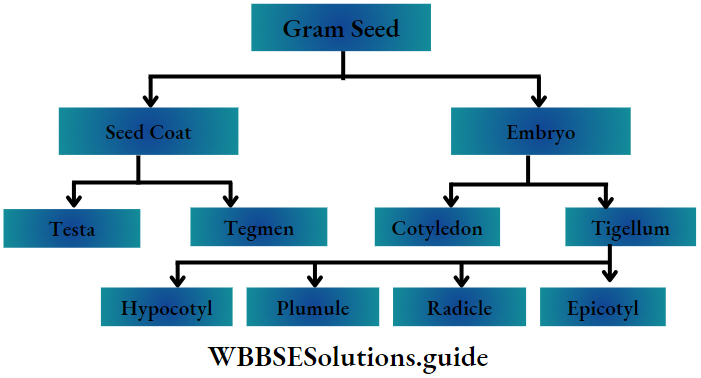
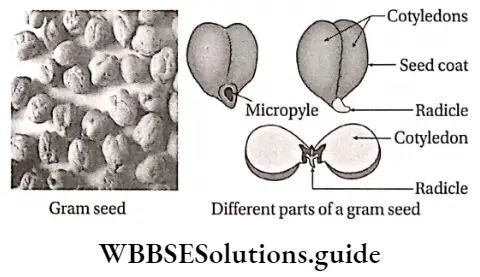
Definition: Monocotyledonous plants are those angiosperms that have a single cotyledon in their seeds.
Monocotyledonous plants General features:
- Generally, they are annual plants, but the trees are perennial.
- Plants are usually herbaceous, with weak stems (for example rice, wheat, grass, etc.). Some may be shrubs and trees (palm, date palm, coconut), while some may be climbers (orchids).
- The embryo contains one cotyledon. The endosperm is well-developed and store food for future use.
- The leaves may be simple or compound. They show parallel venation and sheathing leaf bases.
- Leaves are isobilateral, mesophyll tissue is not differentiated into spongy and palisade parenchyma.
- Stems are generally unbranched, with nodes and internodes. In paddy and bamboo, the internodes are hollow, while in maize and sugarcane, those are solid. In some plants like ginger, onion, etc., stems are underground.
- At the early stage of development, plants lose their tap roots and fibrous root systems develop.
- The flowers are trimerous (the number of whorls present is 3 or in multiples of 3).
- Calyx and corolla fuse to form the perianth.
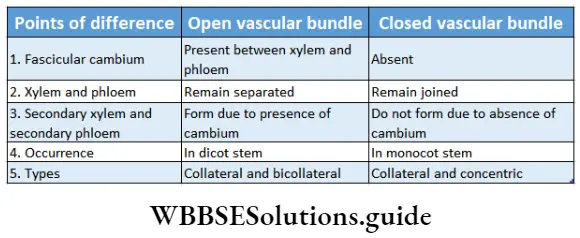
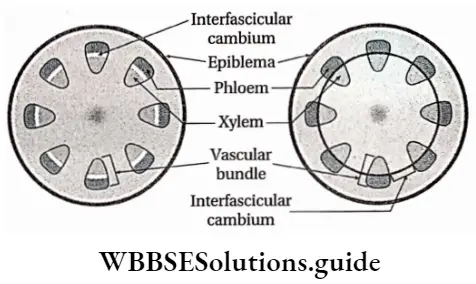
- Stem vascular bundles are conjoint, collateral and closed. The bundles remain scattered throughout the cortex. No stele is formed.
- The number of vascular bundles in the root is usually six or more.
- Due to the lack of cambium in vascular bundles, no secondary growth is observed in the stem and roots.
- The plumule is present near the embryo axis. Coleoptile and coleorhiza are present in seed.
- The seeds show hypogeal germination.
Examples: Pisum sativum, Mangifera indica, Hibiscus rosa-sinensis, etc.
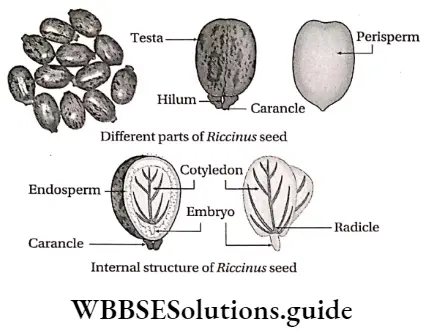
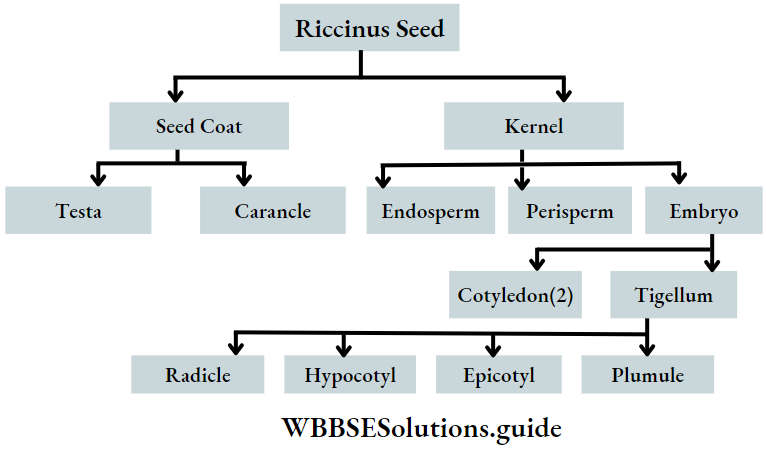
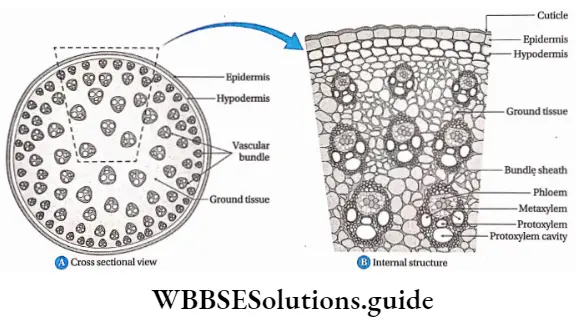
Dicotyledonous Plants
Dicotyledonous plants Definition: Dicotyledonous plants are those angiosperms that have two cotyledons in their seed.
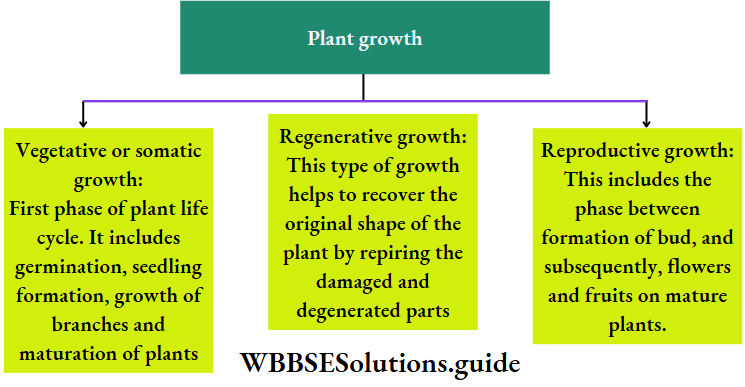
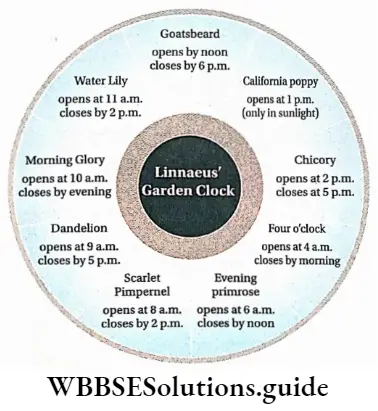
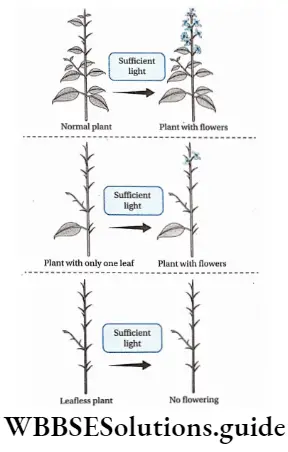
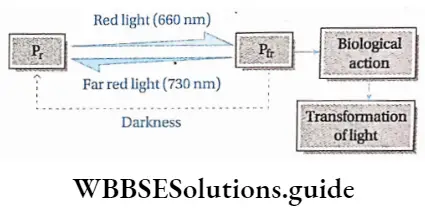
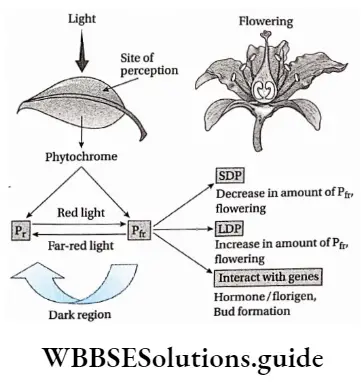
Plant Life Cycles And Alternation Of Generations
A life cycle is a sequence of events that occur from birth to death of an organism. Generation is any entity with distinct morphological features that represent a particular event or phase in the life cycle.
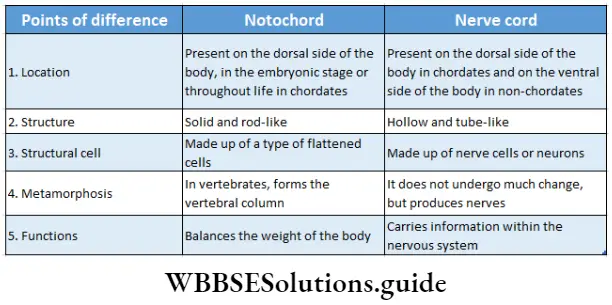
Based on chromosome number an organism can be haploid or diploid. In plants, haploid gametes are produced by gametophytes while the diploid structures that produce spores by meiosis are called sporophytes.
In their life cycles, plants have the gametophytic or sporophytic phase as the dominant one, in which they pass most of their life span. Depending upon these dominant phases, life cycles are mainly of three types.
Those types are discussed below—
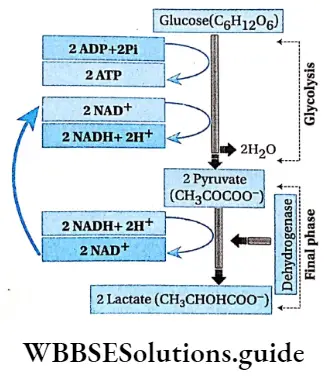
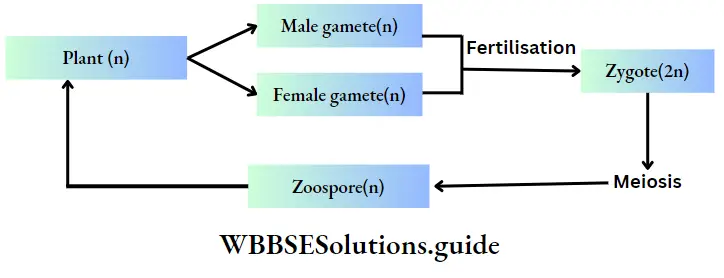
Haplontic or haplobiontic life cycle:
- In this type of life cycle, the gametophyte (haploid) stage is dominant throughout the life cycle and the sporophyte (diploid) stage is represented only by the zygote.
- The gametophytic plant develops haploid gametes inside the gametangia.
- The fusion between these two gametes forms the zygote.
- The zygote immediately undergoes meiotic division and forms four haploid meiospores.
- These meiospores develop into haploid plants. The alternation of generations can be interpreted by changes in the chromosome number.
Example: This type of life cycle is observed in Volvox sp., Chlamydomonus sp., Spirogyra sp., etc.
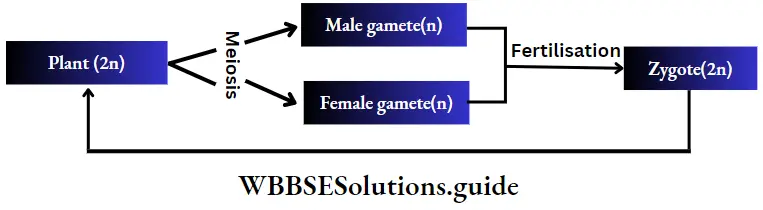
Diplontic or diplobiontic life cycle:
- In this type of life cycle, the plant body is sporophytic and diploid and the haploid gametophytic phase is represented only by the gametes.
- Sporophytic plants bear gametangia inside which gametes are produced by meiosis.
- The gametes upon fertilisation form diploid zygote.
- The zygote gives rise to a new sporophytic plant body.
“plant kingdom taxonomy and classification explained”
Example: This type of life cycle is observed in Cladophora sp., Fucus sp., etc., algae and in all seed-bearing plants i.e., gymnosperms and angiosperms.
Haplo-diplobiontic life cycle:
- In this type of life cycle, both the haploid and diploid phases are prominent.
- They differ in chromosome number and reproductive function. The haploid gametophytic plant reproduces sexually, i.e., by producing gametes.
- The gametes produce diploid zygotes through fertilisation. The zygote gives rise to the diploid sporophyte.
- The sporophytic plant reproduces by asexual processes, i.e., by producing haploid spores.
- In this life cycle, alternation of two generations occurs by sporogenic meiosis and fusion of gametes.
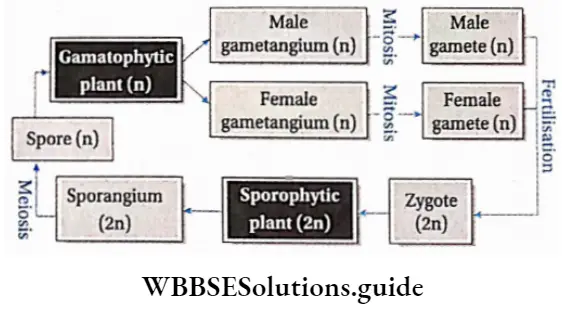
- Isomorphic or homologous types, i.e., gametophyte and sporophyte are morphologically similar. Example: This type of life cycle is observed in Ectocarpus sp., Polysiphonia sp., etc.
- Heteromorphic or heterologous type, i.e., the gametophyte and sporophyte are morphologically dissimilar. Example: This type of life cycle is observed in bryophytes and pteridophytes.
Plant Kingdom Notes
- Alkaloid: A class of nitrogenous organic compounds of plant origin having physiological actions on humans.
- Cilium (plural—cilia): An organelle found in eukaryotic cells. In some organisms, cilia are the means of motility.
- Coenocytic: A multinucleate thallus/body with a continuous mass of protoplasm enclosed by one cell wall.
- Coleoptile: A sheath protecting the tip of the plumule in monocots.
- Coleorhiza: A sheath protecting the tip of a radicle in monocots.
- Dichotomous branching: A pattern of branching where an equal division of an apical bud or apical growth point forms two equal branches.
- Dorsiventral: An organ or body of an organism having dissimilar upper (dorsal) and lower (ventral) surfaces.
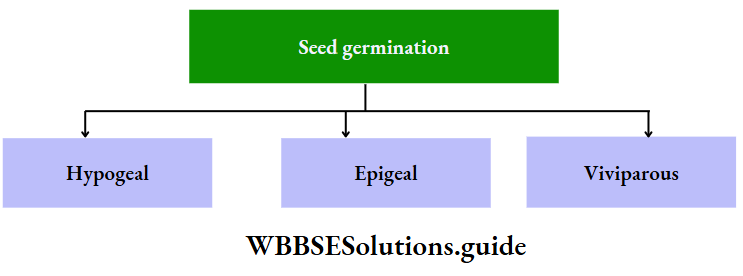
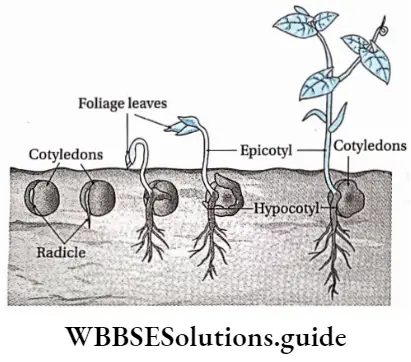
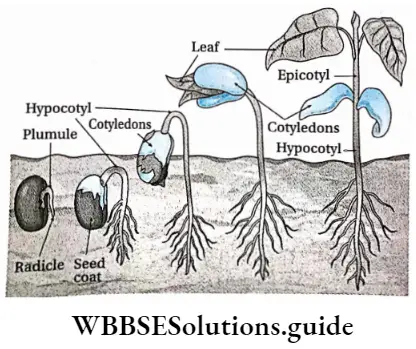
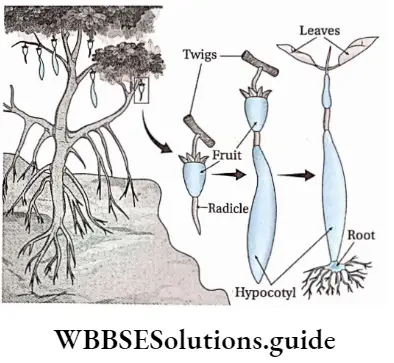
- Epigeal: Germination of seeds with one or more seed leaves or cotyledons appearing above the ground.
- Hypogeal: Germination of seeds with cotyledons appearing above the ground. Plumule: Part of the embryo which will grow into the shoot.
- Radicle: Part of the embryo which will grow into root.
- Sporopollenin: It is an inert biopolymer found in the tough outer wall of spores and pollen grains.
- Tinsel flagellum: A type of flagellum which has fine minute hairs along the axis.
- Whiplash flagellum: A type of flagellum which has a smooth surface.
- Xylan: A type of polysaccharide found in the cell walls of plants and algae.
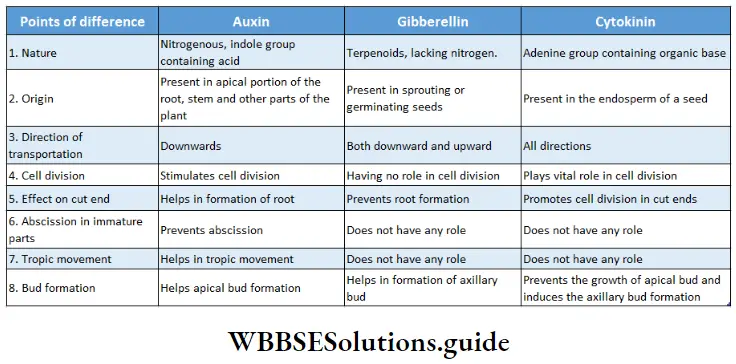
Points To Remember
- Linnaeus first used the term algae.
- The branch of science that deals with the study of algae is called phycology.
- Body of algae is a simple thallus, that consists of parenchymatous cells.
- Large marine algae are known as seaweeds or kelp.
- The longest phase in the life cycle of algae is its gametophytic phase.
- Stored food in green algae are starch and oil.
- Red algae appear reddish due to the presence of the pigment phycoerythrin.
- The pigment responsible for the brownish colour of brown algae is fucoxanthin.
- The main stored food of Phaeophycean algae is laminarin.
- Algae mostly exist as haplobiontic, diplobiontic and triplobiontic.
- The cell wall of Chlamydomonas sp. lacks cellulose.
- Volvox sp. is a freshwater green algae that forms colonies.
- Volvox sp. was first observed by Antonie van Leeuwenhoek in 1700.
- Chlorella sp. is a unicellular green algae that is used as a food.
- Chlorella sp. is a source of antibiotic chlorellin.
- The common name of Spirogyra sp. is pond silk.
- The algae Spirogyra sp. has three types of reproduction—vegetative, asexual and sexual.
- Algae are consumed by various organisms in the aquatic ecosystem.
- lodin is obtained from some phaeophycean algae such as Laminaria digitatia, Ecklonia sp., etc.
“plant kingdom classification PDF notes download”
- Bryophytes are known as amphibians of the plant kingdom.
- Alternation of generations is observed in mosses.
- Although bryophytes are terrestrial, they require water during the process of fertilisation.
- The sporophytic phase of the life cycle in bryophytes depends on the gametophytic plant body for food.
- The common name of bryophytes of the Class Anthocerotopsida is hornworts.
- Apospory can be seen in some species of Anthoceros.
- Seedless, vascular plant fern has approximately 10,000 species.
- Salvinia sp., Azolla sp., and Ceratopteris sp., are aquatic ferns.
- Pteridophytes have a distinct vascular system made up of xylem and phloem tissues. They are called vascular cryptogams.
- Generally, four types of ferns are seen—Psilopsida, Lycopsida (clubmoss or ground pine), Sphenopsida (horsetail) and Filicopsida or Pteropsida (ferns).
- Lycopsids are of two types-(a) homosporous (like Lycopod/t/m sp.) and (b) heterosporous (like Selaginella sp.).
- The aquatic fern Azolla is used as organic manure.
- Lycopodium sp. is used to cure skin diseases.
- Gymnosperms are intermediates between ferns and angiosperms.
- Psilotum sp. is known as a primitive fern. It lacks true roots. Leaves are small and lack veins.
- Sequoia sempervirens is the largest gymnosperm (366 ft), Zamia sp. is the smallest gymnosperm (25 cm), and Sequoiadendron gigantia is the oldest gymnosperm (3,500 years based on the annual ring count).
- The vascular bundle of gymnosperms is conjoint, collateral and open.
- Gnetum sp. is the intermediate between gymnosperm and angiosperm.
- The wood obtained from gymnosperms is called softwood.
- The endosperm of gymnosperms are produced before fertilisation, so the endosperms are haploid (n).
- Polyembryony is observed in gymnosperms, like, Pine.
- The largest ovum, ovule and antherozoa are present in Cycas sp.
- The gymnosperm Ginkgo biloba (maidenhair tree) is known as a living fossil.
- Cycas revoluta is the scientific name of the plant sago palm.
- Pine tree bears numerous pollens which are yellow in colour. These pollens have wing-like structures and are dispersed by air. When these are carried in the air, it appears like a yellow cloud. This is known as a ‘shower of sulphur’ or a ‘shower of golden dust’.
- Fertilisation in pine trees may occur one year after pollination. Polyembryony is seen in pine trees. However, they produce one embryo after maturation.
- Angiosperms are vascular plants, that form fruits, containing seeds.
- Angiosperms are classified into two divisions— monocotyledons and dicotyledons
- Angiosperms are of three types—herbs, shrubs or trees. They can also be annual, biennial or perennial.
“plant kingdom important characteristics and divisions”
- Some species of P|ants on|Y grow other plants but do not obtain food from the host. These are known as epiphytes. Aerial roots of epiphytes absorb water from air through the spongy cells, called velamen, present at the tips.
- Endosperms of angiosperms are produced by double fertilisation (fusion of a male gamete to a diploid secondary nucleus). Thus, endosperm cells have a triploid (3n) set of chromosomes.
- The vascular bundle in the roots of angiosperms is radial and exarch. Number of the vascular bundles is more than 6 in monocots and 2-6 in dicots.
- The vascular bundle in the stem of angiosperms is conjoint, collateral or collateral, open or closed and endarch.
- Secondary growth is seen in dicot plants but it is not seen in monocot plants.
- The largest flower is Rafflesia Arnoldi, whose diameter is 1 meter and weighs approximately 15 kg. The smallest flower and smallest angiosperm is Wolffia microscopic (duck weed).
- Largest angiosperm in Eucalyptus regnance.