
Concept Of Metabolism
Definition: The sum total of all chemical reactions constantly occurring in the living organism, that are necessary for maintaining the living state of cells and that of the organism, is termed as metabolism.
The metabolic reactions take place as a series of linked reactions. These multi-step processes are called metabolic pathways.
Metabolic pathways of a living system can be categorized into two types—
- Anabolic pathways and
- Catabolic pathways.
Anabolic pathways
These pathways are constructive in nature and result in the formation of complex structures from simpler ones. Anabolic pathways consume energy and hence, are endergonic in nature.
” metabolism definition”
For example, amino acids are linked together to synthesize proteins. Protein synthesis requires energy input and is an example of the anabolic pathway.
Catabolic pathways
These pathways are destructive in nature and lead to the breakdown of complex substances into simpler ones.
Catabolic pathways are made up of exergonic reactions because they release energy.
For example, glycolysis is a catabolic pathway where glucose breaks down to produce two molecules of pyruvic acids.
The energy released during catabolic pathways is stored in the form of chemical energy in the high-energy bonds of ATP (Adenosine triphosphate) molecules.
ATP acts as a link between exergonic and endergonic reactions. This is because ATP is hydrolyzed to release the chemical energy needed for various endergonic (energy consuming) reactions.
Living Stage
A living system exists in a steady state, characterized by particular concentrations of all the biomolecules present within the living system.
The steady state is in a non-equilibrium state because the living system is working continuously. It prevents itself from reaching the state of equilibrium so that it is able to perform work.
Types of metabolism in the human body
The rate of formation, utilization, and transformation of biomolecules is called turnover. The rate of turnover of molecules in a metabolic pathway is called metabolic; flux.
For being in the state of metabolic flux, the metabolites tend to reach equilibrium. The equilibrium state is not reached because the end products of the metabolic pathways are utilized again.
| Class 11 Biology | Class 11 Chemistry |
| Class 11 Chemistry | Class 11 Physics |
| Class 11 Biology MCQs | Class 11 Physics MCQs |
| Class 11 Biology | Class 11 Physics Notes |
Thus, the living organism tends to attain a steady state by metabolic flux. This takes place in the presence of energy, which is produced as a result of different metabolic reactions.
So, we can conclude that, without metabolism, a living state cannot exist.

According to Lehninger and others, biomolecules are normally present as essential organic components of living systems, and their essential organic compounds are collectively called metabolites.
These are mainly of two types—
Primary metabolites: They are formed as intermediates and produced via normal metabolic pathways in almost all living organisms, e.g., sugar, amino acids, nucleotides, lipids, proteins, etc.
Secondary metabolites: They are formed especially by the alteration of primary metabolites, mainly in the plant’s body, e.g., alkaloids, pigments, aromatic compounds, terpenoids, etc.
Enzymes
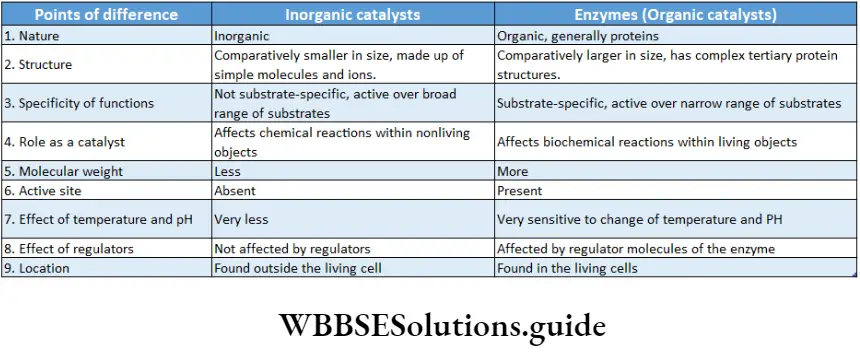
Enzymes Definition: Enzymes are protein molecules acting as biological catalysts that are able to accelerate the rate of chemical reactions in cells. While they remain unchanged at the end of the process.
“what is metabolism “
Enzymes Discovery:
- Jon Jakob Berzilius (1835) was the first to term the activity of enzymes ‘catalytic’.
- Louis Pasteur (1850) concluded from his studies that fermentation of sugar to alcohol is catalyzed by a vital force in living yeast cells, which he termed as ‘ferments’
- The term ‘enzyme’ was coined by W. Kuhne in 1878.
- It is derived from the Greek words, En= in and Zyme = yeast.
- In 1897, Edward Buchner was the first to isolate the enzyme, zymase, which was able to ferment sugar. (f)J.B. Sumner first purified and crystallized the urease enzyme from jack bean.
- After this, John Northrop (1930) isolated many enzymes like ‘pepsin’, and ‘trypsin’ in crystalline form. Sumner and Northrop were awarded the Nobel Prize in Chemistry along with Wendell Stanley in 1946.
- Thomas Cech and Sidney Altman (1983) were awarded the Nobel Prize in chemistry for the discovery of the catalytic property of RNA.
Properties of enzymes: The properties of enzymes are discussed in separate heads below.
Reversibility: Most of the enzymes are able to catalyze reversible reactions.
This property of the enzyme is dependent on various factors. For example, fumarase catalyses the conversion of malic acid to fumaric acid at pH 7.8 and the reverse reaction occurs at pH 6.2.
\(\text { Malic acid } \frac{\text { Fumarase }(\mathrm{pH}: 7.8)}{\text { Fumarase }(\mathrm{pH}: 6.2)} \text { Fumaric acid }\)Reusability: Enzymes remain unchanged and are released after the completion of the biochemical reactions. The unchanged enzyme is reused again.
Solubility: Enzymes are proteinaceous in nature and are soluble in water, mild glycerol, sodium chloride solution, and alcohol.
Proteinaceous nature: Enzymes are generally globular proteins. All the enzymes have a specific amino acid sequence.
Usually, they are colloidal in nature and have high molecular weights. For example, the molecular weight of bacterial ferredoxin is 6000 Da.
Carry charge: Enzymes are charged molecules.
Due to the presence of amino acids, each enzyme has a charge. The charge depends on the pH of the solution.
Biocatalysis: The enzymes show catalytic properties. They cannot initiate or stop a reaction, but only accelerate the rate of reactions. They also remain unchanged at the end of the reaction and, thus, can be reused as minute quantities are required for every reaction.
Buffering capacity: Enzymes have a buffering capacity (acid-base). They are amphoteric molecules that behave both as acids and bases.
Isoelectric point or pH (I): Each enzyme has a specific isoelectric point. It is a specific pH at which the net charge of protein equals zero so that it does not move in an electric field.
pH temperature sensitivity: Due to the proteinaceous nature, enzymes are extremely sensitive to the pH and temperature. Their structure and activity depend on these two factors.
Most enzymes have an optimum pH which ranges between 4 to 9 and the optimum temperature range for the activity of the enzymes is 37°-40°C.
At very low temperatures, the enzymes are not active. At high temperatures of 50°-60, °C usually enzyme activity is usually damaged, however, few enzymes are activated at higher temperatures (e.g., Taq polymerase from Thermus aquaticus).
For most enzymatic reactions, a 10° C rise in temperature doubles the rate of reaction. While the temperature decreases by 10° C the rate becomes half.
This measure of the rate of change of a biological or chemical system as a consequence of the change of temperature by 10° C is called Q10 (temperature coefficient).
\(\begin{array}{r}\mathrm{Q}_{10}=\frac{\text { Rate of the reaction at temperature }(\mathrm{t}+10)^{\circ} \mathrm{C}}{\text { Rate of the reaction at temperature } \mathrm{t}^{\circ} \mathrm{C}} \\
\text { [Mostly, value of } \mathrm{Q}_{10}=2 \text { to 3] }
\end{array}\)
Denaturation: When enzymes are heated or exposed to extremely high temperatures, the non-covalent bonds between peptide chains break giving rise to the primary structure of the enzyme which is a protein.
This unfolding of protein is due to the loss of secondary, tertiary, and quaternary structures. Due to this effect of denaturation, the loss of activity takes place due to the loss of the active site.
“metabolic process “
Specificity: Enzymes catalyze specific reactions. Each enzyme is specific for each reaction.
This catalysis occurs in the special site where the reactants (known as substrates in enzymatic reactions) bind and react to form the products.
This site is known as the active site of the enzyme, which is specific for the substrates. For example, sucrase catalyzes the hydrolysis of sucrose only. Similarly, protease breaks down proteins.
Activity: The activity of the enzyme refers to the rate of reaction at which it binds to the substrate. Enzymes lower the activation energy of the reactions.
The measure of enzyme activity, i.e., specific activity is usually expressed as n moles of substrate transformed to products per minute per mg of the enzyme under optimal conditions of measurements.
It can catalyze the conversion of about 104-105 units of the substrate into products, in one minute.
Substrate concentration: The activity of the enzymes depends on the concentration of the substrate. The activity of the enzymes increases up to a certain concentration of the substrate, after which it attains a maximum velocity.
Location:
- It is present in all living cells. They may remain within the cells in inactive form. These are called proenzymes or zymogens. They are converted to active enzymes only in the presence of certain factors.
- Some of the intercellular enzymes remain within certain organelles such as nucleus, mitochondria, plastid, etc. They may also remain within the cytosol or attached to the plasma membrane, etc.
- Extracellular enzymes like pepsin are synthesized within the cells of some organs but are secreted outside the cells, where they show their action.
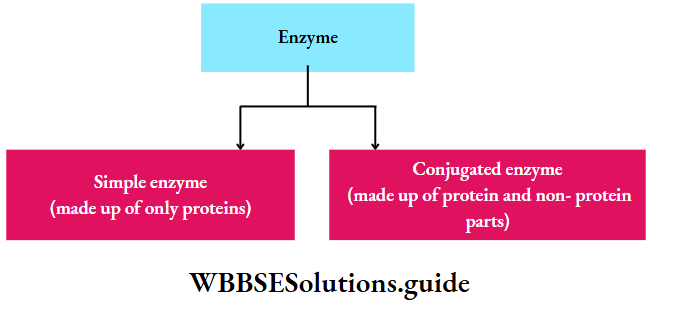
Chemical nature of enzymes: All enzymes are protein in nature but ribozyme is a complex of the ribosomal RNA (rRNA) and protein. This catalyzes the synthesis of protein from the mRNA template. Generally, the enzyme is of two types-
“metabolism definition and example “
Simple enzyme: It comprises only a protein part called apoenzyme. E.g., pepsin, trypsin, amylase.
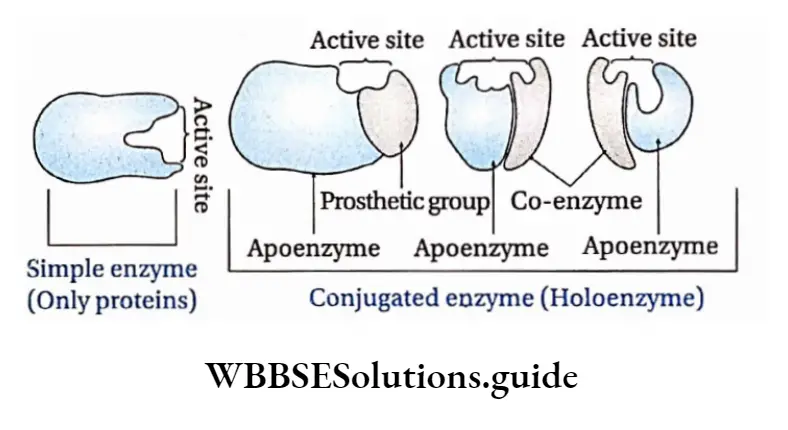
Conjugated enzyme: It comprises a protein part or apoenzyme with a non-protein part or cofactor. A cofactor can be inorganic or organic in nature. Conjugated enzymes are also called holoenzymes.
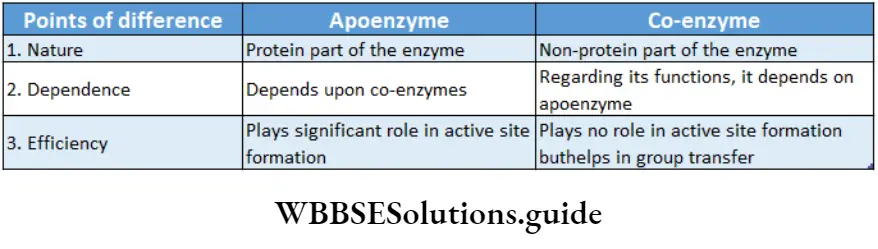
Apoenzyme: The protein part of the enzyme is called apoenzyme. The inactive form of the apoenzyme is known as a proenzyme or zymogen.
The proenzyme may contain several extra amino acids in the protein. These additional amino acids are removed by proteolysis.
Metabolism process step by step
This allows the final specific tertiary structure to be formed before it is activated as an apoenzyme.
The apoenzyme is thermolabile. It is destroyed under high temperatures and on application of heat. The apoenzymes have one or more active sites to which co-factors attach and form the holoenzyme.
“metabolism concept “

Cofactor: The nonprotein part of the enzyme responsible for the catalytic activity is called cofactor. Cofactor is thermostable. There are three recognized categories of cofactors. These are of the following types
Inorganic ion: Inorganic ions are alternately called metal activators. The different types of inorganic ions like Cu2+, K+, Fe2+, Mg2+, Zn2+, Mn2+, etc., play an important role in activating the catalytic reaction. For example, zinc is a cofactor of the enzyme carboxypeptidase.
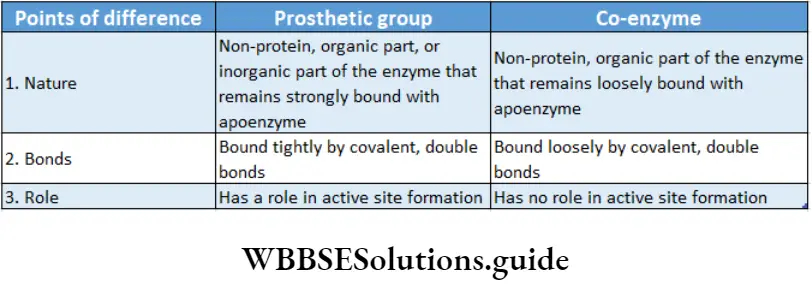
Prosthetic group: The organic nonproteneous cofactor or inorganic factor that remains firmly attached to the protein part of the enzyme through a covalent bond and assists in the catalytic activity, is known as the prosthetic group.
For example, in peroxidase and catalase, which catalyze the breakdown of hydrogen peroxide to water and oxygen, haem is the prosthetic group and it is a part of the active site of the enzyme.
Co-enzyme: The organic non-proteinaceous, heat-stable part of an enzyme that remains loosely attached to the apoenzyme, is called a coenzyme.
Usually, they attach only at the time of catalysis. E.g., NAD (Nicotinamide adenine dinucleotide), NADP (Nicotinamide adenine dinucleotide phosphate).
This is derived from nicotinic acid and can exist in both oxidized and reduced forms.
Metalloenzyme
The enzymes which contain metal ions, that are directly bound to the protein or to enzyme-bound non-protein components, are called metalloenzymes. E.g., Carbonic anhydrase is a zinc metalloenzyme.
Structure of the enzyme: Enzymes are proteins and are made up of long chains of amino acids. The linear chains may fold to form tertiary structures.
Different enzymes have different sequences of amino acids.
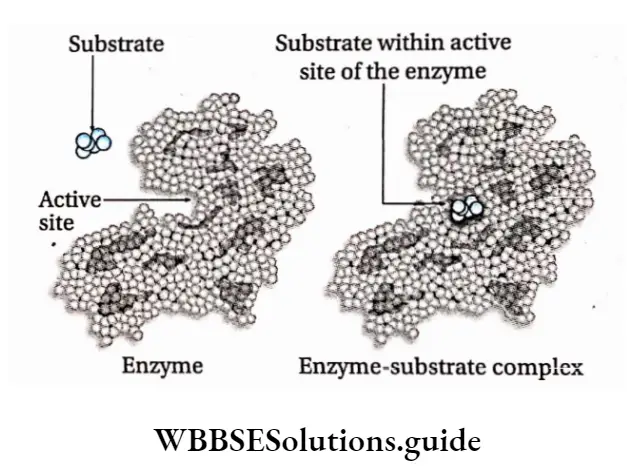
The active site is a location on the enzymes that are formed when the enzymes fold into their functional shape.
The substrate binds to the enzyme at the active site. The complex formed by the union of the enzyme and its substrate is called the enzyme-substrate complex.
Once the substrate has been chemically modified, meaning the reaction ends, it becomes a product.
“metabolic function “
That product then gets released from the active site of the enzyme.
Nomenclature and classification of enzymes
- Enzymes react upon specific substrates. They are named according to the substrate with the suffix -ose or by adding ‘lytic’ to the name of the substrate.
- Enzymes can be named according to the reactions they catalyze. For example, oxidase (this type of enzyme catalyzes oxidation reactions). Enzymes can also be named according to the type of substrate of the reaction they catalyze.
- In such cases, ‘in’ may be added to the name of the substrate. E.g., pepsin, trypsin, etc.
- Enzymes may also be named according to their source. For example, papain is obtained from papaya.
- According to a modern process of naming, enzymes may have two parts in their names.
- The first part denotes the substrate while the second part denotes the reaction catalyzed by the enzyme. Example: glutamate-pyruvate transaminase.
Nomenclature and classification of enzymes based on International rules
- In 1961 the commission of enzymes of the International Glutamic Acid + Pyruvic Acid Union of Biochemistry and Molecular Biology (IUBMB) framed certain rules for the nomenclature and classification of enzymes.
- The commission recognized six different classes of enzymes, which were further divided into subclasses.
- The enzymes are now named according to their systematic position and are given the commission number. In this pattern of naming the first number indicates the class to which the enzyme belongs and subsequent numbers indicate sub-class in that order.
- For example, nitrate reductase, or nitrate oxidoreductase, is named E.C.1.7.1.1. Here, E.C. stands for enzyme commission.
The number 1, denotes the class of enzyme—oxidoreductase.
Here, the number, 7 (1.7) means that it belongs to the subclass of enzymes that acts on another nitrogenous compound as electron donors.
The next number (1.7.1) denotes that the enzyme has NAD+ or NADP+ as an acceptor of electrons.
The last number 1 (1.7.1.1) indicates the above enzyme as the first enzyme of the series. This enzyme helps in the reduction of nitrate to nitrite.
The main six classes of enzymes are discussed under separate heads below.
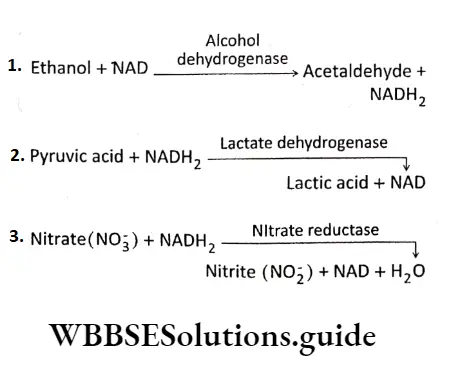
E.C.l or Oxidoreductases: The enzymes that catalyze oxidation and reduction reactions of their substrate, i.e., the enzymes that catalyze the transfer of H or O atoms or electrons from one molecule to another are known as an oxidoreductase.
E.g., Oxidases (cytochrome oxidase), reductase (nitrate reductase), dehydrogenase (lactate dehydrogenase), etc.
“metabolism diagram “
E.C.2 or Transferases: Transferases are the enzymes that catalyze the transfer of a functional group from one molecule to another. E.g., transaminase (leucine Transaminase), Phosphotransferase(2′-Phosphotranferase),etc.

E.C.3 or Hydrolases: The enzymes which catalyze the breaking down of specific bonds of various compounds by the addition of water molecule are known as hydrolases.
In hydrolysis reactions, C-O, C-N, and C-S bonds are cleaved by the addition of H20 in the form of OH- and H+ to the atoms forming the bond. Example Esterase (acetylcholine esterase), glycosidase (protease), peptidase (trypsin), etc.
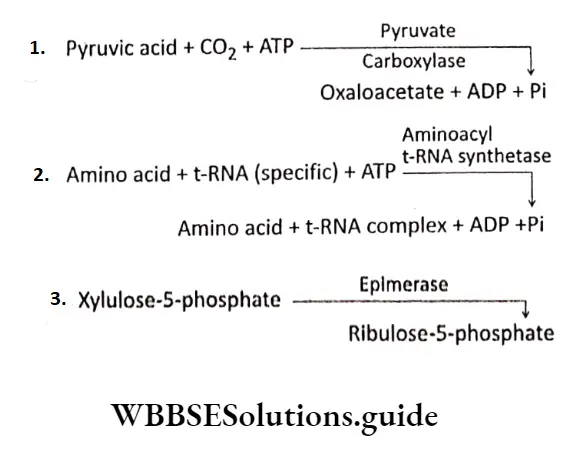
\(\text { Sucrose }+\mathrm{H}_2 \mathrm{O} \longrightarrow \text { Sucrase } \longrightarrow \text { Glucose }+ \text { Fructose }\) \(\text { Protein }+\mathrm{H}_2 \mathrm{O} \stackrel{\text { Protease }}{\longrightarrow} \text { Peptides }\)E.C. 4 or Lyases: Lyases are the enzymes that catalyze the cleaving of C-C, C-O, C-N, and C-S bonds without hydrolysis or oxidation. E.g., decarboxylase (pyruvate decarboxylase), aldolase (fructose bisphosphate aldolase), etc.

E.C.5 or Isomerases: Enzymes that catalyze the isomerization reactions that cause intramolecular rearrangement of atoms in the substrates and thus form one isomer from another, are known as isomerases. Examples are isomerase (phosphohexo isomerase), epimerase (4-hydroxyproline epimerase), etc.

E.C.6 or Ltgasc (Synthetase): Ligases are the enzymes that catalyze the synthesis of C-C, C-S, C-O, and C-N bonds in reactions coupled with the cleavage of high-energy phosphate bonds in ATP or some other nucleotide.
Examples: carboxylase (pyruvate carboxylase), synthetase (glutamine synthetase, aminoacyl tRNA Synthetase), ligase (malate CoA Ligase), Etc.
Mechanism Of Enzyme Action
The action of enzyme depends on different factors. Enzymes are biocatalysts in nature.
All the reactions in a living organism’s body are catalysed by enzymes.
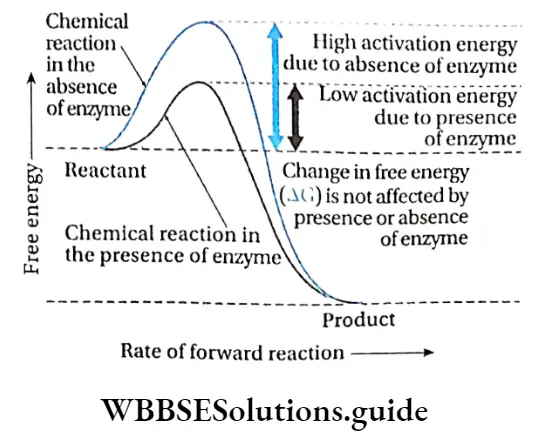
But to begin the reactions, some amount of energy is required to initiate the reaction. Enzymes enhance the rate of chemical reaction by lowering their activation energy.
Activation energy
The activation energy of a reaction is the amount of energy in calories required to bring all the molecules in one mole of a substance at a given temperature to the transition state at the top of the energy barrier.
At this point there is equal probability for reactants to undergo reaction to form products or to fall back and remain unreacted.
Michaelis and Menten described the activity of the enzymes with respect to activation energy. They observed that reactions that are not catalysed by enzymes, occur at a slower rate and require higher activation energy.
For example, hydrolysis of casein requires 20600 kcal/mol in the absence of any enzyme, while only 12600 kcal/mol is required in the presence of the enzyme.
Collision theory
According to this theory, the rate of a chemical reaction is proportional to the number of collisions between reactant molecules.
So, the more often the reactant molecules collide, the more often they react to each other and the faster is the reaction rate.
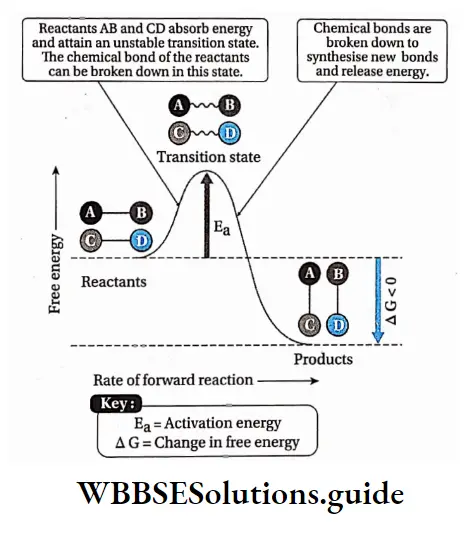
The point in the chemical reaction where there is the maximum value of energy is called transition state.
The reactant molecules while colliding with each other, if they hit with enough energy to go through the transition state, they will react and form new molecules.
At the transition state, new bonds are formed while old ones are broken.
Models Of Enzyme Action
Two models have been proposed regarding the mode of enzyme action:
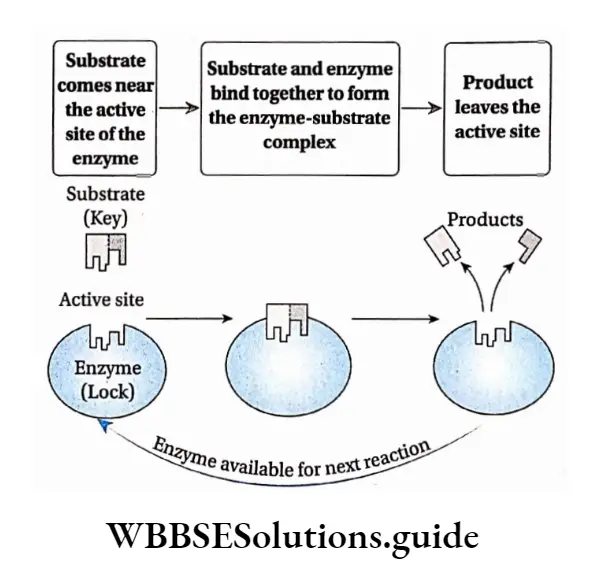
Lock and Key hypothesis: According to this hypothesis, the enzyme active site has configuration complementary to the shape of the substrate so that the enzyme and substrate can recognise and fit to each other. This model was proposed by Fischer (1890).
According to this model Each enzyme [E] has a specific active site or catalytic site into which only the substrate [S] fits.
The enzyme [E] and the substrate [S] both possess complementary conformation, which fits exactly into one another just like a key fits into a lock.
The active site of the enzyme is rigid and fixed, which further helps in the binding.
After binding, the enzyme [E] and the substrate [S] form the ES complex.
This [ES] complex immediately breaks down to the product [P] and enzyme [E] and product is released from catalytic site of the enzyme.
Difference between catabolism and anabolism
\(\mathrm{E}+\mathrm{S} \rightleftharpoons \mathrm{ES} \longrightarrow \mathrm{P}+\mathrm{E}\)Special groups like -NH2, -COOH etc., help in binding the enzyme and the substrate. Co-enzymes and activators may also influence the binding.
Limitations of the model: Fischer explained enzyme specificity and rigidity of the binding sites of the enzymes.
But he could not explain the allosteric behaviour of enzymes (binding to a molecule at a site other than the active site) which emphasises that the active site of enzyme changes its conformational ensemble in order to bind with the substrate.
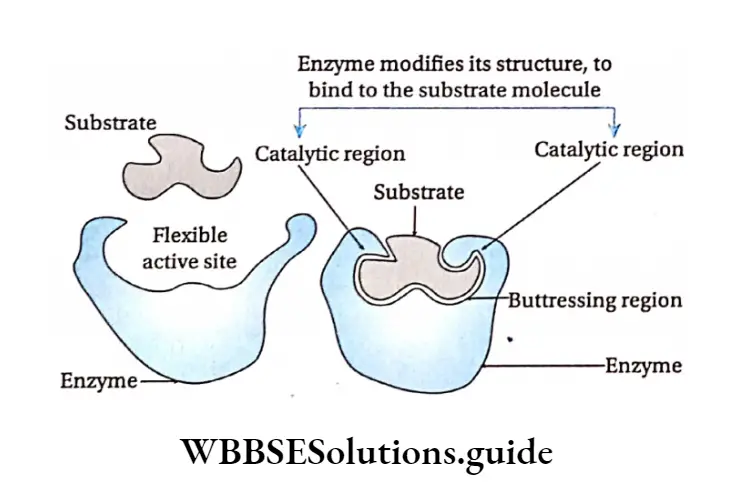
Induced fit hypothesis: Koshland et al. (1959) suggested another model of enzyme action with some modifications to the lock and key hypothesis.
According to this model the active site contains two groups—buttressing region and catalytic region.
But tressing region binds the substrate and forms a complex. Catalytic region weakens the bond of the substrate by electrophilic and nucleophilic processes and leads to the formation of the product maximum velocity or Vmax
This hypothesis proposes that, the enzymes and their active sites are not rigid, rather physically more flexible.
When a substrate combines with the buttressing region of an enzyme, it induces conformational changes in the enzyme structure.
This brings the catalytic region opposite to those bonds of the substrate that are to be weakened. The amino acids in the active site are modified into specific structure.
This enables the enzyme to bind with the substrate and perform its catalytic activity effectively.
The substrate changes into product and gets released from the buttressing site due to structural changes in the site. As a result the product is released.
Regulation Of Enzyme Action
The action Of enzymes are regulated by different factors.
Factors affecting enzyme action
Substrate concentration: The catalytic activity of enzyme is influenced by substrate concentration.
The rate of enzyme action increases with increasing substrate concentration.
If the substrate concentration is increased gradually from a very low concentration, the enzyme activity at first rises proportionately.
If this is plotted on a graph it will show a steep rise in the initial velocity (V0) With further increase in the substrate concentration, the initial rise in velocity of enzyme action falls and ultimately reaches a point where this velocity neither increases nor decreases. This point is called the maximum velocity or Vmax.
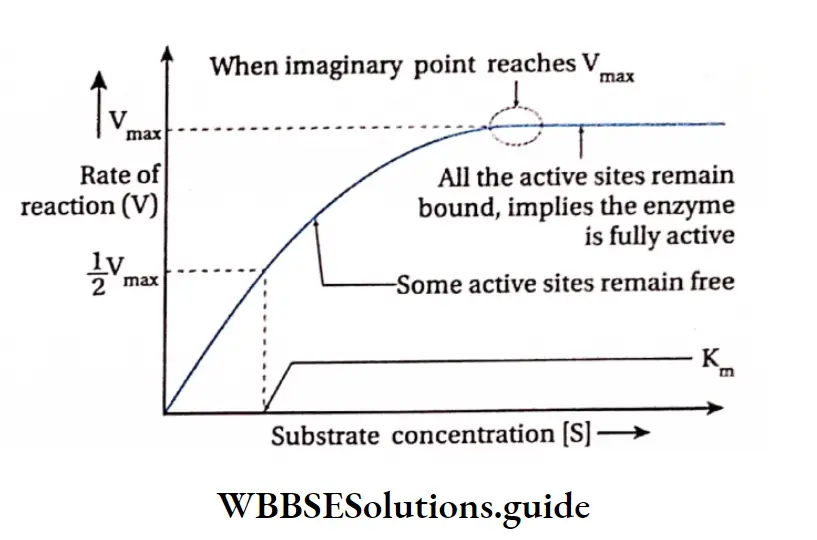
The initial reaction rate, V0 can be described by Michaelis-Menten equation as proposed by Leonor Michaelis and Maud Menten (1930).
\(\mathrm{V}_0=\frac{\mathrm{V}_{\max }[\mathrm{S}]}{\mathrm{K}_{\mathrm{M}}+[\mathrm{S}]}\)where, Vmax= Maximum velocity
S = Substrate concentration
KM = Michaelis-Menten constant
V0 = Initial velocity
The substrate concentration at which half the maximum velocity, -y is attained by an enzyme catalysed reaction is called KM or Michaelis-Menten constant.
When initial velocity (V0) of an enzyme catalysed reaction is half of the maximum velocity (vmax) i.e., V0 \(v_0=\frac{v_{\max }}{2},\) hen the equation changes as follows—
\(\text { When, } v_0=\frac{v_{\text {max }}}{2}\) \(\frac{V_{\max }}{2}=\frac{V_{\max }[S]}{K_M+[S]}\) \(\text { or, } \quad \frac{1}{2}=\frac{[S]}{K_M+[S]} \text { or, } K_M+[S]=2[S]\)or, Km =[s]
Thus the Michaelis-Menten constant [KM] is the amount of substrate concentration [S] when the initial velocity of an enzyme action is half of the maximum velocity \(\left[\frac{V_{\max }}{2}\right]\).
The KM value is an indicator of the affinity that an enzyme has for its substrate. Therefore, the value of KM is different for different enzymes. The KM value of the protease, which acts on a variety of proteins, varies with the type of protein.
An equation with a high KM indicates that the enzyme does not bind efficiently with the substrate, and Vmax will only be reached if the substrate concentration is high enough to saturate the enzyme.
Temperature: Enzymatic reactions usually take place between 20°-40°C, but at high temperature of 60’-70°C they get denatured as they are thermolabile. Generally the enzymatic action is maximum at an optimum temperature of 37C.
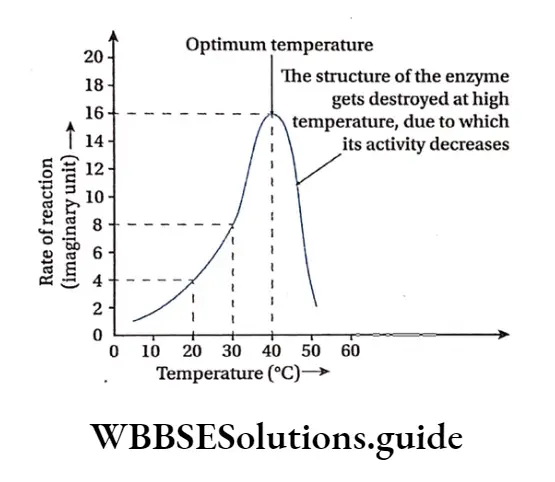
Enzyme concentration: The increase in the concentration of enzyme increases the rate of enzyme action within the cell.
At constant temperature and pH, a high level of substrate concentration increases the rate of enzyme action proportionately.
Product concentration: A high concentration of accumulated product inhibits an enzyme to catalyse a forward reaction.
This type of inhibition is known as feedback inhibition or product inhibition.
pH: Enzymatic action is sensitive to pH. Each enzyme shows its maximum activity at a particular pH known as optimum pH. Some enzymes are active in acidic medium while others in alkaline medium. Each enzyme can act efficiently at its optimum pH. E.g., pepsin acts at an optimum pH of about 2.0 while trypsin acts at an optimum pH of 8.5
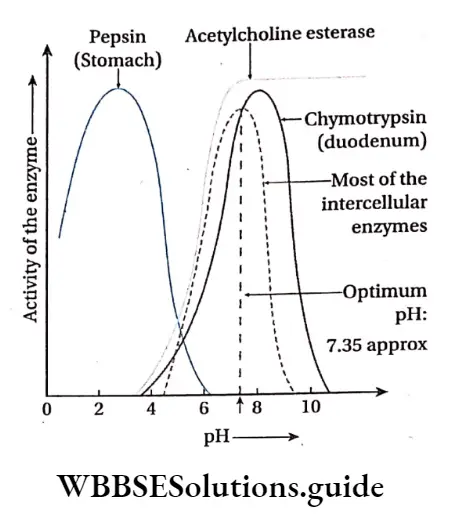
Methods of regulation of enzyme action
Enzyme action can be accelerated or inhibited by allosteric modulation and feedback regulation.
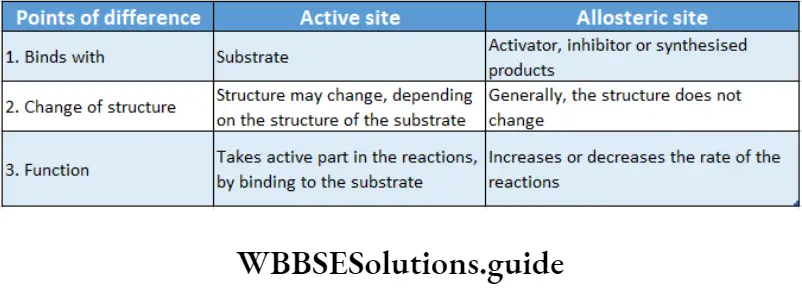
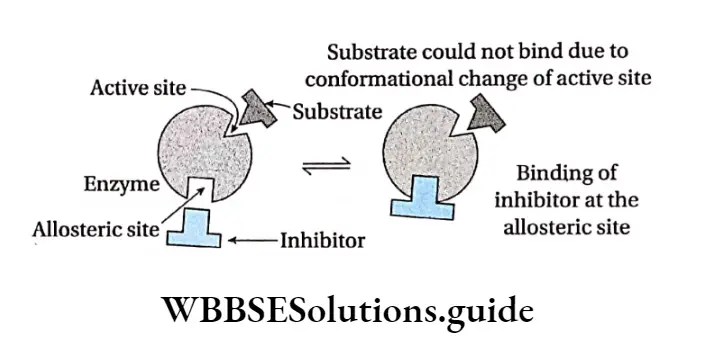
Allosteric modulation: The property of an enzyme by which an effector molecule binds to any site other than the active site of the enzyme, resulting in acceleration or inhibition of enzyme action is known as allosteric modulation.
Allosteric site: Some enzymes possess a second site of activity other than the active site, called the allosteric site. Such enzymes are known as allosteric enzymes.
Allosteric enzymes do not obey Michaelis-Menten kinetics. The effector molecule or allosteric modulator binds to the effector or allosteric site while the substrate molecule binds to the active site for the reaction to take place.
Allosterism: Some modulator molecules upon binding to the allosteric sites of the enzymes, influence the enzyme activity. This phenomenon is called allosterism.
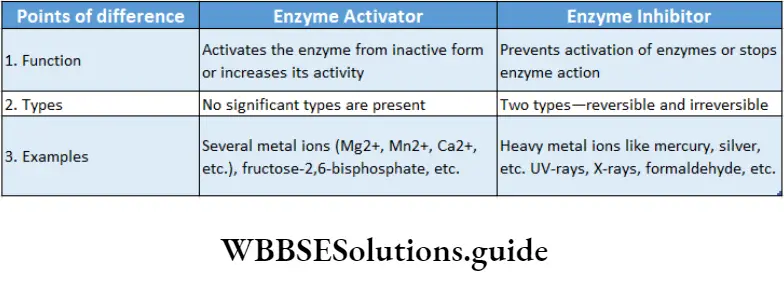
Compounds which can speed up the enzyme action are known as allosteric activators or inducers while those which slow down the reaction rate of an allosteric enzyme are known as allosteric inhibitors.
Types of allosterism: According to the property there are two types of allosterism—
Positive allosterism: When an effector or inducer molecule that binds with the allosteric site of an enzyme, enhances the binding capacity or the functional efficacy of the active site, then it is known as positive allosterism. In this case, the reaction proceeds in the forward direction resulting in formation of product.
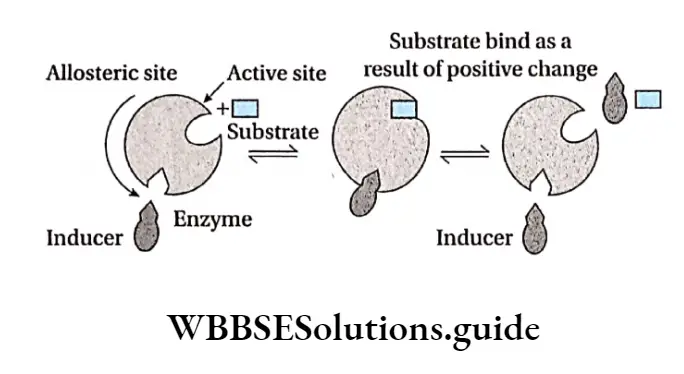
This in turn increases the enzyme activity and hence the rate of the reaction increases. An example of positive allosterism is the binding of oxygen molecules to haemoglobin.
Negative allosterism: Sometimes, when an inhibitor molecule binds with the allosteric site of an enzyme, the conformation of the active site changes. As a result, the substrate cannot bind to it. This is known as negative allosterism.
In this type, the reaction is inhibited so, product is not formed. This in turn, reduces the rate of the reaction. For example, when 2,3-bisphosphoglyceric acid binds to an allosteric site of haemoglobin, the affinity towards oxygen decreases in all subunits of the protein.

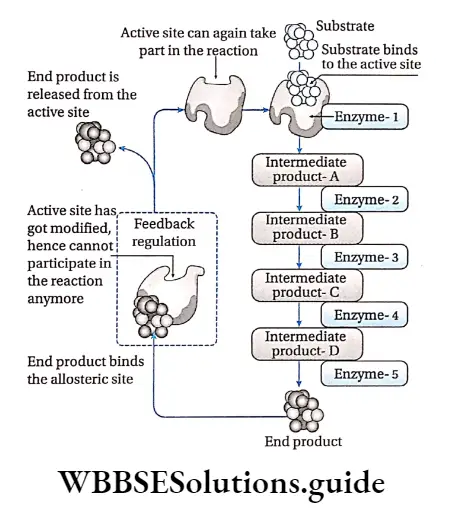
Feedback regulation: When the concentration of the end product of a series of enzymatic reactions regulates the activity of the enzymes involved, the phenomenon is called feedback regulation.
When large amount of product is produced, it binds to an allosteric site of the first enzyme in that reaction series, thus, inhibiting its activity. When the concentration of the end product decreases, it frees the allosteric site and the enzyme reverses back to its active state.
For example, the amino acid isoleucine is synthesised through a series of biochemical reactions from the amino acid threonine. An adequate amount of isoleucine as an end product in the cell, inhibits the activity of the enzyme threonine deaminase.
It is caused by feedback inhibition which prevents the further synthesis of isoleucine. A decrease in the concentration of isoleucine in the cell, activates the threonine deaminase enzyme by feedback mechanism.
Enzyme Inhibitors And Enzyme Inhibition
The substances that inhibit the activity of the enzymes are called enzyme inhibitors and the mechanism by which they do so is called enzyme inhibition.
Types Of Enzyme Inhibition
Enzyme inhibition is of two types—
- Irreversible and
- Reversible.
Irreversible inhibition: The method of inhibition by which the activity of the enzymes is inhibited permanently and cannot be restored, is called irreversible inhibition.
Characteristics: An irreversible inhibitor binds to the active site and changes its structure permanently or destroys the protein structure of an enzyme.
It stops the enzymatic activity and the enzyme cannot be recovered.
Heavy metal ions, SH groups, radiations like UV rays, X-rays, are responsible for irreversible inhibition.
For example, salts of cyanides inhibit the activity of cytochrome oxidase essential for cellular respiration, resulting in cell death.
Reversible inhibition: The method of inhibition by which the activity of an enzyme is inhibited temporarily is called reversible inhibition.
Characteristics: In this type of inhibition, some inhibitor molecules may attach to the active site by non-covalent interactions.
It may also involve feedback inhibition or modification of the active site by positive modulator, etc.
When these inhibitors are released from the active site, the enzyme reverses back to the functional state.
Types: There are three types of reversible inhibition—competitive, non-competitive and uncompetitive.
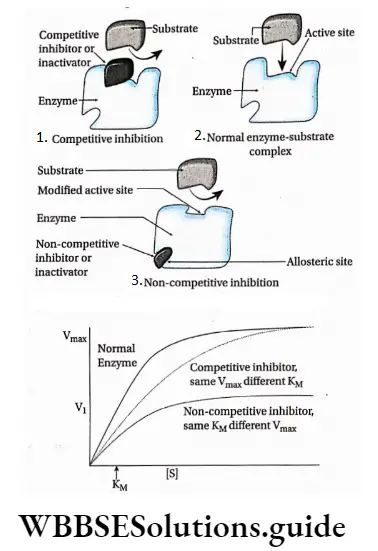
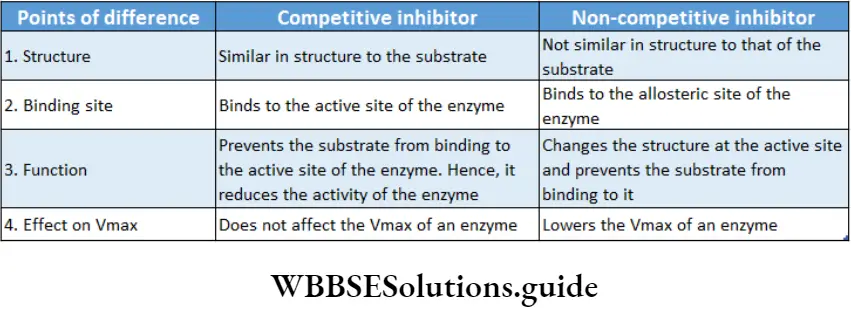
Competitive inhibition: Competitive inhibitors are molecules that are structurally similar to the substrates and compete with the substrates to bind at active sites.
This leads to slowing down of the reactions. The mechanism is called competitive inhibition. Competitive inhibitors increase KM value, thus decrease substrate affinity of the active site. Vmax remains unchanged.
Examples of metabolism in everyday life
For example, malonate is the competitive inhibitor of succinate dehydrogenase. Malonate binds to the active site of succinate dehydrogenase, preventing succinate from binding to it.
Non-competitive inhibition: Non-competitive inhibitors have no structural similarity with the substrate but may form an inhibitor-enzyme complex at the allosteric site of the enzyme.
It inactivates an enzyme by changing the structure of the active site. Thus, the substrate can no longer recognise or bind to the active site of the enzyme.
This prevents the enzyme-substrate complex formation. Non-competitive inhibitors lower Vmax of an enzyme but keeps the value of KM unchanged.
For example, non-nucleoside reverse transcriptase inhibitors or NNRTIs (like Etravirine) can bind to the reverse transcriptase enzyme of HIV, inhibiting the synthesis of viral RNA without interfering with substrate binding. This inhibitor is used as an anti-HIV drug.
Uncompetitive inhibition: Uncompetitive inhibitors bind to the enzyme after formation of enzyme-substrate complex.
It binds close to the active site. This stops the action of the enzyme by altering the structure of enzyme-substrate complex.
As a result, KM decreases. The Vmax also decreases as a result of removal of activated complexes.
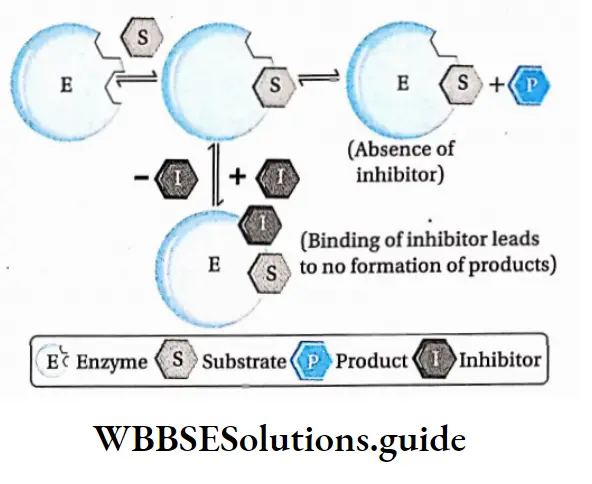
Significance Of Enzyme Inhibition
- Excess products can be regulated by feedback inhibition.
- An idea about the different metabolic reactions within the body can be derived.
- Structure and activity of the enzymes can also be explained by these theories.
- Different medicines and pesticides can be designed applying the knowledge of enzyme inhibition.
Denaturation of enzymes
High temperature, high energy radiation (UV rays, X-rays etc.), heavy metals (Ag+, Hg 2+, As+), salts, concentrated acids, alkali etc., affect the active site of the enzymes. This in turn changes the structure of the proteins, breaking the hydrogen bonds. This finally destroys the enzyme integrity permanently.
Isoenzyme or Isozyme
- The different forms of an enzyme, differing only in certain amino add sequences but catalysing the same reaction, are called isoenzymes.
- Generally, different isoenzymes are found within different cells and tissues.
- Due to slight difference of the molecular structure, they have different properties (like favourable pH, affinity towards substrate, effect of inhibition etc.).
- For example, lactate dehydrogenase enzyme of human cells, has five isoenzymes.
- An enzyme, a-amylase, of wheat endosperm has 16 isoenzymes. Alcohol dehydrogenase of maize has 4 isoenzymes.
Allozyme
Allozymes or alloenzymes are variant forms of an enzyme which are coded by different genes of same locus. E.g., DNA polymerase.
Proenzymes or Zymogens
Proenzymes or zymogens are biologically inactive substances which are metabolized into enzymes. For example, HCl of gastric juice alters the inactive I pepsinogen to active pepsin enzyme.